
This is a continuation of my blog 'coverage' of our new monograph - for an introduction to the big new paper, see the first post in this series here. For the third post in this series, click here.
But first - it's been a pretty dark day for me and Sarah, so I need to share some memes that were made out of/inspired by my artwork:

Family portrait of Coronodon that I illustrated last year and finished in February: upper left, juvenile Coronodon havensteini; lower left, adult Coronodon havensteini; upper right, Coronodon planifrons; lower right, Coronodon newtonorum. Graphite and digital, 2023. Original about 10x14".

My illustration has inspired some pretty fantastic memes - my artwork does look like the other Coronodon whales are laughing at the one. Cartoon by M. Nur Yahya on twitter.

A variation of an older meme - by @natespithe on Twitter.

One of my favorites - by Henry Tsai on Facebook.

Stratigraphic Distribution of Coronodon
I'll dive into the rationale for naming new species shortly, but first I'd like to discuss the stratigraphic distribution of specimens. The holotype skull of Coronodon havensteini was collected from a submerged exposure of the Ashley Formation - a late early Oligocene sandy limestone. It's firm, with some concretions here and there, generally kind of a brownish tan to pale yellow color and occasionally can be olive green or khaki - frequently fossiliferous, with sparse mollusks, barnacles, and bryozoans. The Ashley Formation has been dated with Strontium isotopes to about 28-30 Ma - uppermost Rupelian, which is the stage occupying the first half of the Oligocene epoch. All new specimens we referred to Coronodon havensteini are also from the Ashley Formation. The two specimens that ended up being holotypes of new species - CCNHM 166 and ChM PV 2778 - were both collected from the Chandler Bridge Formation. When Al Sanders and others named the Chandler Bridge Formation in 1986, they remarked that the whale faunas were so similar that they were likely only separated in time by about a million years or so. Somewhat surprisingly, strontium isotope dates indicate that there's about 4-5 million years - and that the Chandler Bridge Formation is 24.7-23.5 Ma or so (Weems et al., 2016). Point is - these Coronodon specimens are separated by a bit of time. There are some other subtle differences: eomysticetids seem to be stratigraphically separated, with all specimens of Micromysticetus being derived from the Ashley and all specimens of Eomysticetus originating from the Chandler Bridge. Xenorophid odontocetes appear somewhat stratigraphically separated: more plesiomorphic taxa, like Xenorophus and Albertocetus, make up most of the xenorophid assemblage from the Ashley whereas there are additional, more derived taxa in the Chandler Bridge including Cotylocara and Echovenator, as well as at least one additional Cotylocara-like genus that has not yet been named. There are perhaps a half dozen waipatiids in the Chandler Bridge, but only two or three forms from the Ashley, and they seem to be restricted: a dwarf form, and Ediscetus osbornei named a few years ago. Another species of Ediscetus may be present in the Ashley - and that glut of waipatiid specimens will be my major project this summer.

Stratigraphic distribution of Coronodonidae in the Oligocene rocks of Charleston, South Carolina.

The holotype skull of Coronodon planifrons, CCNHM 166, the Volcko-Chapman whale.
Recognizing the Existence of New Species of Coronodon
Discovering a new species - whether you're digging it up and doing all the sweaty work, or the actual anatomist doing the descriptive work - is not always obvious. For a long time I thought there was a single species - after all, I've already reported (or interpreted) both of the odontocetes Albertocetus meffordorum and Ankylorhiza tiedemani as being present in the Ashley and Chandler Bridge formations. My working hypothesis was "one species in both units" - which I feel, from the perspective of a lumper, the most conservative approach. When I arrived at CCNHM in 2015, the specimen that is now the holotype of Coronodon planifrons, the "Volcko whale" CCNHM 166, and the "Coosaw whale" CCNHM 164, were both in our collections but not included in the original study. The "Volcko whale" was found in 2010 and the Coosaw whale a few years earlier (each within a couple miles of our house!). I sat down and looked for differences earlier on back in 2018 and actually failed to make a number of observations I made later - but I did make one important observation back then: the holotype specimen of Coronodon havensteini, and what is now the holotype of Coronodon planifrons (CCNHM 166), had differently shaped supraorbital processes of the frontal. There are these little processes on either side of the eye socket, helpfully named the preorbital and postorbital process (measuring between them, by the way, gives you eye socket size). In Coronodon havensteini these processes are the same thickness if you measure from the inside out. However, in CCNHM 166, the postorbital process is quite a bit thicker than the preorbital process (and vice versa). Now, these bones are also very porous so I thought, maybe this is just caused by wear to the skull, and discounted species-level differences.

A comparison of the preorbital and postorbital processes of the three species of Coronodon - some of the earliest observations I made. Coronodon havensteini has pre- and postorbital processes of equal depth, Coronodon planifrons has a deeper postorbital process, and Coronodon newtonorum has a deeper preorbital process.
About a year ago I noticed that the supraorbital process of the frontal, in anterior view, was nearly horizontal - and I only noticed this because I but the CCNHM 166 skull, along with referred C. havensteini specimen CCNHM 164, backwards, with the posterior end of the skull facing the wall. In both CCNHM 164 and the holotype CCNHM 108, the supraorbital processes descend at an angle and are decidedly not horizontal. I think I made that observation during a meeting. So, I pulled the specimens out and started more detailed comparisons. I noticed that the sagittal crest was much longer in CCNHM 166, and that the frontal-parietal suture is much more V-shaped. I also noticed that the sternomastoid fossa - a D-shaped rugose pit for the sternomastoid muscle, which is positioned on the squamosal bone and just above the jaw joint - has this deep trough that rises up along the side of the nuchal crest; this is missing in Coronodon havensteini. At this point I was more or less convinced that the number of anatomical differences had exceeded individual variation for Coronodon havensteini and instead suggested to me that CCNHM 166 represented a different species.

A parallel development happened in Fall 2017 after the first paper was published - collector Taffie Chapman, who found parts of CCNHM 166 in the field, donated specimens she had first found and kept. A bunch of missing parts clicked back together - for example, the condyle of the mandible was in her material! Taffie kept one tooth for herself, a nice premolar, and I did ask for it - but she politely insisted, and I knew better than to push it. Included amongst her material were several more vertebrae and a bunch of partial teeth. As a result, we now call it the "Volcko-Chapman whale". Years later, last summer actually, Jonathan Geisler and I finally got a chance to compare these side by side and we realized several teeth from the original sample were improperly glued together - and, within 20 minutes of when Geisler was supposed to drive back to New York after a vacation here in Charleston. I airscribed away some putty in the prep lab and texted him photos of the preliminarily reassembled teeth - he texted back something along the lines of "this changes quite a bit."

The periotics (ear bones) and dentition of Coronodon planifrons, the Volcko-Chapman whale.
One of the first changes were the smallest (and furthest posterior) molars - the smallest one had broken in half, one half found and kept by each collector. I glued them back together, revealing a tiny upper third molar (M3) that was approximately 2/3 the diameter of the second to last tooth (upper second molar, M2). The M3 is unknown in the holotype, but the upper M2 is preserved, and the "Coosaw whale" specimen of Coronodon havensteini (CCNHM 164) has both the M2 and M3. These teeth are of similar size in Coronodon havensteini, and also larger than both teeth in CCNHM 166 (Coronodon planifrons). In other words, CCNHM 166 "Volcko-Chapman whale" (Coronodon planifrons) has 1) smaller molars and 2) molars that decrease in size posteriorly. We initially thought there were some differences in cusp counts, but that changed when we settled upon the final identification of teeth to position after discovering the tiny size of the M3 - after this, the cusp counts were quite similar. I took another look at the earbones, and found that there were also a few features of the periotic bone that were unique - for example, the lateral tuberosity extends far laterally, whereas it is a small bump in the holotype of Coronodon havensteini. All of these features ended up in our diagnosis* of Coronodon planifrons. The tiny size of the upper M3 also permitted us to refer an isolated molar to Coronodon planifrons, also showcasing what the tooth shape would be without any tooth wear. I suggested naming the species after its most distinctive feature: horizontal frontal bones over the orbit, hence Coronodon planifrons.
*The first time I read a scientific paper and saw diagnosis for a species I thought "what the hell? A diagnosis is for a disease" but as it turns out, a diagnosis in zoology and botany (or taxonomy in general) is a list of the different anatomical/morphological features that are used to identify specimens of that species. In a sense, it is one of the most important parts of the reporting of a new species, genus, or family. Ewan Fordyce impressed upon me how important it is to craft a useful, lasting diagnosis that future researchers can use. One pet peeve of mine is the lazy use of synapomorphies from a cladistic analysis and restricting your diagnosis to just features from the cladistic character list: these diagnoses are usually not very useful. Medical diagnoses are constructed similarly: a suite of diagnostic features used to identify a medical condition.
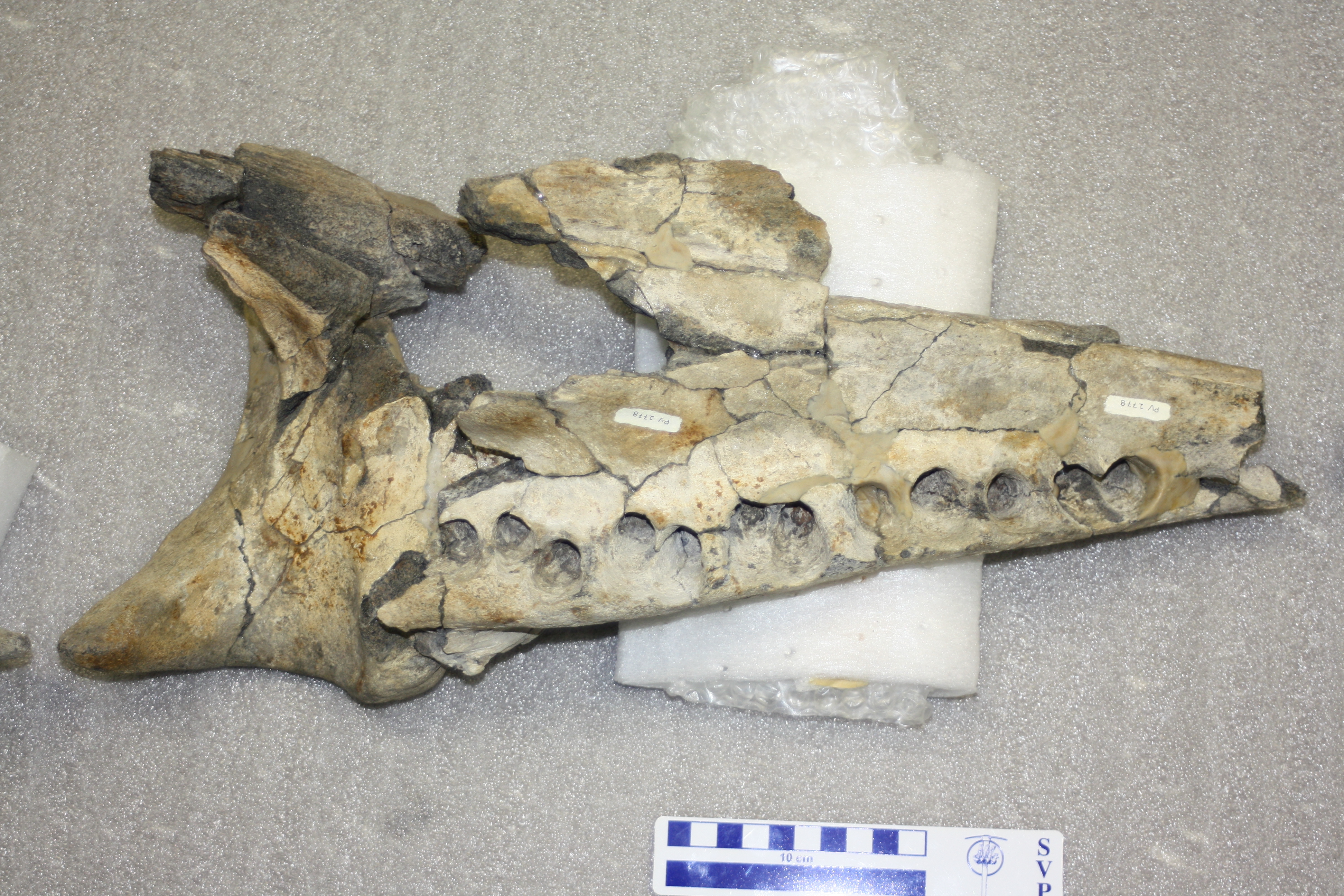
The main skull part of ChM PV 2778, the holotype of Coronodon newtonorum - as I first saw it in 2012.

Placing the mandible of ChM PV 2778 (Coronodon newtonorum) back into place for the first time.
In parallel, I had not looked at ChM PV 2778 - the "Newton" whale - for quite some time. I knew it had a partial mandible and much of a skull, and some earbones. I looked at it again in early spring 2022 and had the following revelations: 1) the mandible could actually be pieced back together, and it was in three pieces until I asked curator Matt Gibson to reconstruct it; 2) there was a nice isolated cheek tooth bearing the same number that at some point had been separated and was stored in a different cabinet. After the first revelation (which, mind you, was about the same time we realized that Al Sanders' conference abstracts indicated this specimen, and "Hoss" - ChM PV 5720 - had 12 instead of 11 mandibular teeth), I took a couple snapshots of the mandible fragments reassembled (not yet glued) and realized it looked quite a bit different from Coronodon havensteini. Instead of a straight ventral margin, the ventral margin in the "Newton whale" (ChM PV 2778) was convex. Likewise, the alveolar (dorsal) margin of the mandible was more strongly concave than in Coronodon havensteini. I looked again at the rostrum, and it too had a convex margin - indicating that ChM PV 2778 had a "smiling" profile of the lower jaw. Out of caution, we waited for Matt Gibson to glue it back together in case I was wrong - but it looked the same when I returned to take photos for the paper. This was the first time I thought that there might be a third species present - which surprised me: I assumed that we might have two chronospecies: C. havensteini in the late early Oligocene Ashley Formation and a younger chronospecies in the Chandler Bridge. I assumed this out of being cautious, conservative, and being a lumper. I swear to god, some folks - and dinosaur researchers in particular are guilty of this sin - seem to think every specimen is its own species and genus.

The holotype skull, teeth, and mandible of Coronodon newtonorum, ChM PV 2778, the Newton whale.
Upon further examination, I noted that the isolated cheek tooth of ChM PV 2778 is an upper tooth, and that it fit best into the sockets (alveolus) for the upper M1. The tooth is somewhat smaller than the M1 of Coronodon havensteini, an observation I duplicated for the lower 2nd premolar. The periotic is quite a bit more slender than in the holotypes of Coronodon havensteini or Coronodon planifrons - and in many respects, it resembles the juvenile specimens of Coronodon havensteini, despite itself being an adult specimen. One feature it shares with Coronodon planifrons is that long lateral tuberosity of the periotic - something lacking in Coronodon havensteini. As it happens, this specimen had a manuscript name and its own genus name - at this point, known only to those who visit Charleston Museum collections and look at the specimen labels where they're written - but the species name actually was proposed to be "newtoni" by Al Sanders and Larry Barnes. We went with this for two reasons: 1) it honored the earlier work by Al Sanders; 2) Claude and Albert Newton found and collected the specimen; and 3) the Newton family participated in 'volunteer night' at Charleston Museum for many years. There was just one catch, which we discovered during the proofing stage: "newtoni" honors only a single male member of the Newton family... and we didn't know which one, because Claude and Albert Newton both participated in the excavation. So, at the last minute, we changed the name to "newtonorum" to honor the whole family, and Claude and Albert in particular. I have not yet met anyone from the Newton family, but I'm hoping to now that the word is out!

Line drawings of the skulls of Coronodon havensteini, Coronodon planifrons, and Coronodon newtonorum from the Oligocene rocks of Charleston, SC.
Other Specimens and the Importance of Publishing on the the "Dregs"
In addition to describing the new specimens of Coronodon havensteini and the two new species, I also wanted to take the opportunity to include a bunch of more fragmentary specimens. These are specimens that 1) could have conceivably been published as a separate paper but 2) the submission/revision process would have probably eaten up more time than the specimens are worth. Since my time is at a premium compared to earlier in my career, I decided instead to include these as 'accessory' data, so to speak. These come in two categories: 1) a bunch of isolated teeth and 2) a partial cranium from the Cooper River.

The isolated teeth constitute every single specimen we are aware of in museum collections at CCNHM and Charleston Museum. There may be more teeth in the South Carolina State Museum, which we did not sample. The lion's share of these lack good data and are identified as Coronodon sp., since it is unclear what species they belong to. One tooth, mentioned above, was identified as the distinctive M3 of Coronodon planifrons. Confusingly, this specimen was found at the same time and locality as the Coronodon planifrons holotype - but is from the same side as the M3, rather than the opposite side - so must represent a different specimen. The preservation is also not terribly similar. Altogether, there are 13 isolated teeth, found as early as 1973 (and possibly earlier). In contrast, I see at least a couple dozen teeth of the giant dolphin Ankylorhiza collected from the Charleston area and posted onto fossil groups on Facebook every year. I've estimated in the past that there are probably in excess of 100 Ankylorhiza teeth for every isolated Coronodon tooth. I'm only aware of three such teeth found since I started in 2015, and about 2-3 additional teeth remaining in private collections, in addition to the 13 isolated teeth we described. Coronodon is a very rare fossil, and I'm confident we probably have published virtually all of those discovered in the Charleston area.

Partial skull CCNHM 8745 - a partial braincase indicating the presence of a third member of the Coronodon clade, now known as the family Coronodonidae.
A partial skull consisting of a phosphatized braincase with a bit of attached matrix has been floating around in our collection for years - sadly, the collector data is lacking, and the most likely collector - Stephen Miller - passed away a few years ago. According to Mark Havenstein, the preservation resembled fossils from the bottom of the Cooper and Edisto rivers, but not from the Wando River where the holotype of Coronodon havensteini and Inermorostrum xenops were discovered - river fossils there were typically grey to black, rather than brown. A clue to the origin of this specimen came from a sea turtle beak published in 2017 by Rob Weems and Mace Brown, who at the time, noted that the specimen had been collected from the Cooper River near Goose Creek, South Carolina. They identified it as the sea turtle Euclastes, and noted that fossils of the archaeocete Dorudon serratus were found with it on the river bottom and attributed to uppermost Eocene strata. The supposed archaeocete specimens, I was told on a different occasion, were this skull and an atlas vertebra thought to be associated with it - however, the atlas was actually from the giant dolphin Ankylorhiza. The Ankylorhiza atlas suggested Oligocene age - and indeed, there are not really any Eocene rocks in the subsurface there - not as shallow as the river bottom - and the river erodes instead into the Ashley Formation. While it's possible the specimens could have been transported from upstream, the vertebra is clearly from a large odontocete and the skull was a good match for Coronodon or something similar. Further, adhering matrix most closely resembles the Ashley Formation and there are these distinctive little convex clams preserved that are common in Ashley Formation nodules from Lowcountry rivers. I used to think that perhaps it was intermediate in morphology between Coronodon and Basilosauridae, but the more I looked at it the more it resembled Coronodon. The differences are mostly around the interorbital region of the skull: it has tiny, short nasal bones that are flat. Some aspects resemble Coronodon more than Basilosauridae and indicate this specimen belongs to Neoceti - the premaxilla contacts the frontal bone (in Basilosauridae the premaxilla terminates further anteriorly and is 'pinched' out by the nasal and maxilla), and the nasals are quite short with the 'blowhole' positioned far posteriorly, just somewhat in front of the eye sockets (resembling Coronodon) rather than halfway out on the rostrum (like Basilosauridae).
So, why publish on the "dregs" of collections? Two major reasons. First, out of scientific thoroughness: I've seen the dangers of "headhunting" - focusing on only the very best specimens, relegating the "dregs" of museum collections to oblivion. This is an underappreciated problem - from my perspective, I've seen so many specimens forgotten by disinterested or unthorough researchers sit untouched in museum collections. Why should we bother keeping such specimens if they're not going to be useful? I have the radical idea that *most* specimens should be published, as many, if not most, are more informative than is generally appreciated. Second, such specimens can provide unexpected benefits. For example: our suite of teeth sugests that Coronodon is quite rare as far as Oligocene marine mammal fosils are concerned. And, more critically, the 'ugly duckling' skull CCNHM 8745 may not be the most informative specimen of our study, but it does indicate that there are three coronodonid genera present in Oligocene rocks here: Coronodon, the larger Basilosaurus-sized cousin "Hoss" (e.g. ChM PV 5720), and the taxon represented by CCNHM 8745.
Introducing the New Family Coronodonidae
The recognition of at least one other unnamed genus of Coronodon-like whale in this paper - along with a third containing the unstudied specimen ChM PV 5720, aka "Hoss" - led us to name the family level clade containing Coronodon. Unfortunately, Coronodontidae is preoccupied by some Devonian sharks, the type genus of which is Coronodus. Family names in taxonomy are pretty rigid: they have to have "-idae" onto the end of the type genus name. So, the other family I'm familiar with, Eomysticetidae? "-idae" is swapped for the last syllable in Eomysticetus. The correct familial emendation of Coronodon would be Coronodontidae - which is preoccupied. So, in lieu of that, according to the International Code of Zoological Nomenclature (ICZN), you just add "-idae" onto the end of the genus name, however it is spelled. Hence, Coronodonidae - which admittedly does not slip off the tongue quite as nicely as Coronodontidae, but there's nothing I can really do about it. Whenever "Hoss" gets described, it will be named within the Coronodonidae - and is already recognized as one in our paper. This family, by the way, is diagnosed by a long list of features including a blade-like part of the premaxilla that extends in front of the incisors, dorsally curving nasal bones, a lightly articulated (and possibly flexible) rostrum, deep zygomatic processes with enormous sternomastoid fossae just behind them, large cheek teeth bearing at least five mesial cusps, and cheek teeth that are highly emergent from their sockets - among a bunch of other features.
Next up: part 3 - new observations of the growth and functional anatomy of Coronodon and the surprising results of our phylogenetic analysis: is Coronodon actually a mysticete or could it be a late-surviving archaeocete?
6 comments:
Hey Robert, good afternoon. My name is Aretha Guimaraes and I'm a scientist from Brazil. I'm currently working in the Brazilian Amazon and Ive been reading your blog for a while.
Could you please do an update post about your lab notebooks? I find your note taking system very interesting and I'd like to see more about it.
Thanks for your amazing content and documentation work. I really enjoy coming here and learning about paleontology (even though I work with plants, still find it quite interesting).
Best wishes,
Aretha Guimarães
Ola Aretha! I'm glad you're enjoying my occasional writing. What in particular about my notebooks would you like me to update? Field notes, or the way in which I write my specimen descriptions out by hand, diagrams, photo labeling, etc?
Sometimes I forget how much reach this blog has and then I get a comment from someone like you! Many thanks =]
Hey Robert, thanks for answering my comment!
I have a few questions regarding the organization of your notes.
1) I’ve read that you divide your notebooks using tabs according to the specimen that you are describing. But how do you continue your note about a certain fossil if the pages that you left for that description are over? Do you continue in another notebook? And if you do continue to write in another notebook, how do you store the complete description later?
2) By now you should have lots of notebooks stored. Do you keep a “master index” where you can check when you are looking for a particular description or info?
3) Do you make any copies of these notebooks? Like a digital copy or something?
4) I have the same problem that you described in one of your posts, where you told us you had to move to the US with a bag full of heavy notebooks. I’ve been moving a lot recently (Brazil – UK – Amazon and so on) and I’ll probably move a lot until I can become a full professor somewhere. I tried to use a digital notebook in order to be more practical to carry things around (since I’m not available to afford shipping all of my notebooks) but I didn’t like it. Did you manage to solve that problem?
5) Regarding the fieldnotes, do you have a field notebook or do you keep your notes in the same notebooks that you use every day?
6) Are you still using A4 notebooks? When you have a drawing of something larger to stick in your notebook, how you do to prevent damage?
7) Do you keep daily notes like research diary?
Also, I’d like to tell you that I always share your notebook posts with my students so that I can convince them to keep their notes, especially my husband (who is also a scientist) and refused to take notes on stuff. I used your notebooks post to convince him of my point and now he keeps things more organized.
Thanks again for your content! I found you by randomly searching in google a few years ago and I always keep an eye on your posts.
Best wishes,
Aretha Guimaraes
Aretha: I wanted to comment to let you know that I have seen your list of queries, and I will write a new post at some point addressing these questions! Probably sometime next month.
Hello! We thoroughly enjoy your work. We found a piece of jaw with a small tooth that looks like these Coronadon molars but smaller, about 1/4” wide. Could this be juvenile?
Hello Patrick! Please send me a photo (can't post photos to blogger comments) at my email address - which is my last name AT gmail.com. I'd be more than happy to take a look at it!
Post a Comment