
Well, here we are - my tenth year in a row summarizing the year's advances in marine mammal paleontology. I don't comment on all of the new papers, there's just too many - but you'll see some commentary below on selected papers.
2022 had a rough start for me, and I caught Covid in June after a snorkeling trip in the Caribbean; nearly had long Covid. This was a slightly better year publication wise - I had a handful of papers come out, three of them qualified for this year's review. More critically, I finished three legacy projects, all of which are in review: a manuscript just submitted on a new species of the toothless walrus Valenictus from the Purisima Formation near Santa Cruz, a longer monographic description of the toothed baleen whale Coronodon, including description of two new species, and most critically - I completed additions and revisions to a massive review of the vertebrate fossil assemblage from the Purisima Formation of California, first submitted in 1989 by my coauthor Frank Perry, and then slowly updated over the past 14 years by yours truly (yes, I was shocked: Frank invited me to updated it in 2008!). I've got several other manuscripts on early cetaceans (Oligocene) on the way, along with a paper on a fossil sea otter, and a big-ass manuscript on the early dolphin Xenorophus that is hitting the 2/3 completion mark. I also reviewed a ton of papers this year - about 15 - and out of the 68 marine mammal paleontology papers I was not an author on, I reviewed 9 (marked with an asterisk) - about 13% of them!
Only a few archaeocete papers came out this year, but among these, a new genus was named for "Platyosphys aithai" and a new partial skeleton referred to this new genus, Antaecetus. A long-awaited redescription of the late-surviving archaeocete Kekenodon onamata from New Zealand was published. Some new mysticetes have been named - including the toothed mysticete Kaaucetus and the crown mysticetes Persufflatius (a very Dr. Seuss-ish name), Jobancetus, and a new species of the balaenopterid Archaebalaenoptera from Peru. Odontocete papers typically reported fragmentary or referred specimens, but the new sperm whale Angelocetus was named and many new specimens of the platanistoid Notocetus were reported. It was a very good year for delphinids - the new Pseudorca-relative Rododelphis was named from the Pleistocene of Rhodes, and a modern desription of the Pliocene early killer whale Orcinus citoniensis was also published. A few new pinniped papers came out, but mostly report relatively fragmentary remains. Early sirenians received some critical attention, with some more followup studies on the surprisingly well-preserved early dugongid-like sea cow Sobrarbesiren from Spain, and new genus of protosirenid Dakhlasiren named from Morocco.
Regardless of these studies, the real winner this year were big data-rich evolutionary analyses, especially focusing on 3D morphometrics of the skull based on a recently assembled dataset of cetacean skull scans using modern and extinct cetaceans, including Bennion et al., Coombs et al., and Goswami et al. Other analytical studies making a splash this year include Kiel et al., examining the relationship between marine mammal evolution and biogeography with the decline of nautiloids; a new and seemingly exhaustive phylogenetic and biogeographic analysis of sea cows by Heritage and Seiffert; an analysis of cranial telescoping in cetaceans by Buono and Vlachos; and a study of convergent dental adaptations in marine amniotes by Fischer et al.
The usual disclaimers apply:
1) If you think I am missing an article, double check the post from last year. If it was published "online early" in 2020, and just has a formal 2021 publication date, then it counts for 2020. Please do not ask me to shift them from one post to the other - don't be daft.*
2) I have copied the authors (citation style) and the title, as well as the URL; I don't have time to include these all in a complete reference format. Please don't ask me to do that. Don't be daft.*
3) In order to qualify the paper MUST consider fossils, because this is a paleontology list. Marine mammal paleontologists frequently publish on the morphology of extant species. That's not paleontology, though. If a phylogenetic study doesn't incorporate fossil data, it also doesn't count.
4) None of the genus/species names are italicized. This is because I pasted directly into notepad to remove formatting. I will not be italicizing these names so please do not ask me to. Don't be daft.*
*Yes, these are all things [daft] randos have asked me to do in the past. I don't get it. If you want these things, do it yourself.

Aguirre-Fernandez et al. 2022. First records of extinct kentriodontid and squalodelphinid dolphins from the Upper Marine Molasse (Burdigalian age) of Switzerland and a reappraisal of the Swiss cetacean fauna.*
The Swiss Upper Marine Molasse (OMM) documents a transgression event dated to around 21 to 17 million years in which dolphin and other vertebrate remains have been reported. We revised the whole cetacean (whales and dolphins) OMM assemblage available in main collections, focusing on the identification and interpretation of periotics (bone that contains the inner ear). Periotics are rare, but they provide the richest taxonomic information in the sample and hint to environmental associations. Micro-computerized tomography allowed the reconstruction of bony labyrinths for comparisons and environmental interpretations. Three families are represented by periotics: Kentriodontidae, Squalodelphinidae and Physeteridae. The cetacean taxonomic composition of the Swiss OMM reinforces biogeographical patterns reported for the Mediterranean and Paratethys during the Burdigalian at a regional scale and the Calvert cetacean fauna of the northwest Atlantic at oceanic scale.
Comments: An important study highlighting how useful and informative isolated odontocete earbones are. I reviewed this last Januaryish, and had been thinking about a similar approach to a survey of isolated periotics from spoils and beaches from the South Carolina lowcountry. This study gave me just the bit of resolve I needed to start!

The fossil record of marine mammals from the upper Pliocene of the western coast of South America is poorly known, hindering our knowledge about how and when marine mammal faunas attained their modern distribution in the area. Here, we describe a new marine mammal assemblage from the Horcón Formation (upper Pliocene), located in the Valparaíso Region of central Chile. The material reported comprises postcranial remains of an indeterminate seal (Phocidae), Delphinoidea vertebrae, and mysticete elements (still in the field). The Phocidae remains constitute the youngest pre-Pleistocene record of this family in South America, providing new information regarding the timing for the establishment of the modern diversity and distribution of pinnipeds in the Southern Hemisphere. Furthermore, the pinniped and cetacean remains reported indicate that the marine mammal assemblage from the upper Pliocene along central Chile was composed of a mixture of modern and extinct taxa.

Shark–cetacean trophic interactions, preserved as bite marks in the fossil record, mostly correspond to isolated or fragmentary findings that bear limited information about major trophic patterns or roles. Here, we provide evidence of focalized foraging by sharks in the form of tooth bite marks over physeteroids fossil bones from the late Miocene of Peru. These findings indicate that sharks were targeting the forehead of coeval physeteroids to actively feed on their lipid-rich nasal complexes. Miocene physeteroids displayed a broad diversity, including giant predatorial forms, small benthic foragers and suction feeders. Like their extant relatives, these animals exhibited enlarged fatty forehead organs responsible for their sound production capabilities, thus evolving taxon-specific cranial architecture. Bite marks are found on the cranial bones where these structures were attached, indicating that sharks actively targeted this region; but also, in areas that would only be accessible following the consumption of the surrounding soft tissues. The shape of the bite marks and their distribution suggests a series of consecutive scavenging events by individuals of different shark species. Similar bite patterns can be recognized on other Miocene physeteroids fossils from across the globe, suggesting that sharks actively exploited physeteroid carcasses as fat sources.

The repeated return of tetrapods to aquatic life provides some of the best-known examples of convergent evolution. One comparison that has received relatively little focus is that of mosasaurids (a group of Late Cretaceous squamates) and archaic cetaceans (the ancestors of modern whales and dolphins), both of which show high levels of craniodental disparity, similar initial trends in locomotory evolution, and global distributions. Here we investigate convergence in skull ecomorphology during the initial aquatic radiations of these groups. A series of functionally informative ratios were calculated from 38 species, with ordination techniques used to reconstruct patterns of functional ecomorphospace occupation. The earliest fully aquatic members of each clade occupied different regions of ecomorphospace, with basilosaurids and early russellosaurines exhibiting marked differences in cranial functional morphology. Subsequent ecomorphological trajectories notably diverge: mosasaurids radiated across ecomorphospace with no clear pattern and numerous reversals, whereas cetaceans notably evolved toward shallower, more elongated snouts, perhaps as an adaptation for capturing smaller prey. Incomplete convergence between the two groups is present among megapredatory and longirostrine forms, suggesting stronger selection on cranial function in these two ecomorphologies. Our study highlights both the similarities and divergences in craniodental evolutionary trajectories between archaic cetaceans and mosasaurids, with convergences transcending their deeply divergent phylogenetic affinities.
Comments: Rebecca Bennion was our very last research visitors at the Mace Brown Museum of Natural History before the pandemic. She scanned CCNHM 103, the referred skull of Ankylorhiza tiedemani - it's very satisfying to see it in her analyses and figures!

Bianucciet al. 2022. The origins of the killer whale ecomorph.
The killer whale (Orcinus orca) and false killer whale (Pseudorca crassidens) are the only extant cetaceans that hunt other marine mammals, with pods of the former routinely preying on baleen whales >10 m in length and the latter being known to take other delphinids. Fossil evidence for the origins of this feeding behavior is wanting, although molecular phylogenies indicate that it evolved independently in the two lineages. We describe a new extinct representative of the killer whale ecomorph, Rododelphis stamatiadisi, based on a partial skeleton from the Pleistocene of Rhodes (Greece). Five otoliths of the bathypelagic blue whiting Micromesistius poutassou are associated with the holotype, providing unexpected evidence of its last meal. The evolutionary relationships of R. stamatiadisi and the convergent evolution of killer whale-like features were explored through a broad-ranging phylogenetic analysis that recovered R. stamatiadisi as the closest relative of P. crassidens and O. orca as the only living representative of a once diverse clade. Within the clade of Orca and kin, key features implicated in extant killer whale feeding, such as body size, tooth size, and tooth count, evolved in a stepwise manner. The tooth wear in Rododelphis and an extinct species of Orcinus (O. citoniensis) are consistent with a fish-based diet, supporting an exaptative Pleistocene origin for marine mammal hunting in both lineages. If correct, predation by the ancestors of Pseudorca and Orca did not play a significant role in the evolution of baleen whale gigantism.

The East Pisco Basin is one of the forearc basins that formed during the Cenozoic along the coast of Peru due to the subduction of the Farrallon-Nazca plate beneath the South American plate. The sedimentary fi ll of this basin is extensively exposed along the coastal Ica Desert, and includes a succession of Eocene to Pliocene marine sediments that account for a ~50-myr-long history of semi-continuous deposition. These rocks are characterized by an outstanding fossil content that remarkably contributed to our understanding of the evolutionary history of the main groups of Cenozoic marine vertebrates. In the Ica desert, the most common and signifi cant vertebrate remains belong to cetaceans. Knowledge on the fossil cetaceans of the East Pisco Basin has grown dramatically in the last fi fteen years thanks to several international research projects involving, among many others, the authors of the present article. These research eff orts have led to the discovery of several hundred fossil skeletons, the most signifi cant of which have been collected, prepared and partly published. Furthermore, interdisciplinary studies were also conducted in order to provide a high resolution chronostratigraphic framework for this fossil record. Remarkable cetacean specimens come from the Yumaque member of the Paracas Formation (middle to late Eocene), the Otuma Formation (late Eocene), the Lower Miocene (~19-18 Ma) portion of the Chilcatay Formation, and allomembers P0 (14.8-12.4 Ma), P1 (9.5-8.5 Ma) and P2 (8.4-6.7 Ma) of the Middle Miocene to Pliocene Pisco Formation. The Lutetian (42.6 Ma) Yumaque strata are home to the quadrupedal protocetid archaeocete Peregocetus pacifi cus, which documents the fi rst arrival of cetaceans in the Pacifi c Ocean. Geologically younger (36.4 Ma) Yumaque deposits have yielded the holotype skeleton of Mystacodon selenesis, the oldest mysticete ever found. This ancestor of the modern baleen whales had a skull provided with a complete dentition and retained hindlimbs, albeit reduced in size. In the Otuma Formation, a nine-m-long basilosaurid (Cynthiacetus peruvianus) has been discovered. The Chilcatay Formation records the fi rst great radiation of the odontocetes, represented by Inticetidae (Inticetus vertizi), basal Platanidelphidi (Ensidelphis riveroi), Squalodelphinidae (Furcacetus fl exirostrum, Huaridelphis raimondii, Macrosqualodelphis ukupachai and Notocetus vanbenedeni), Platanistidae (aff . Araeodelphis), Physeteroidea (Rhaphicetus valenciae and cf. Diaphorocetus), Chilcacetus cavirhinus, indeterminate Eurinodelphinidae, and Kentriodontidae (Kentriodon). Overall, this roughly coeval assemblage displays a considerable disparity in terms of skull shape and body size that is possibly related to the development of diff erent trophic strategies, ranging e.g., from suction to raptorial feeding. In the Pisco Formation, starting from P0, the baleen-bearing whales (Chaeomysticeti) represent the most frequent cetacean fossils (only a few mysticetes are known from the Chilcatay strata). Two chaeomysticete lineages are found in the Pisco Formation: Cetotheriidae (from Tiucetus rosae in P0 to Piscobalaena nana in P2) and Balaenopteroidea (from Pelocetus in P0 to several undescribed species of Balaenopteridae in P2, testifying to a progressive trend toward gigantism). Odontocetes are rare in P0, the “kentriodontid” Incacetus broggii being the only species described from these strata, but they become more abundant and diverse in P1 and P2. In P1, the commonest toothed whale is Messapicetus gregarius, a member of Ziphiidae featuring an extremely elongated rostrum and a complete set of functional teeth. Another ziphiid from P1 is Chimuziphius coloradensis, known only from the fragmentary holotype cranium. The P1 strata also record the appearance of the crown Delphinida, with the superfamily Inioidea being represented by two small pontoporiids (Brachydelphis mazeasi and Samaydelphis chacaltanae) and one iniid (Brujadelphis ankylorostris). Moreover, P1 is also home to the stem physeteroid Livyitan melvillei; featuring a three-m-long skull and teeth reaching 36 cm in length, L. melvillei was one of the largest raptorial predators and, possibly, the biggest tetrapod bite ever found. Acrophyseter is another macroraptorial sperm whale, distinctly smaller than L. melvillei, known from both P1 and P2. Even smaller in size are the kogiids Platyscaphokogia landinii and Scaphokogia cochlearis, both of which are known from the upper strata of P2. The same allomember is also home to the ziphiids Chavinziphius maxillocristatus and Nazcacetus urbinai, the “kentriodontids” Atocetus iquensis and Belenodelphis peruanus, and undescribed members of Phocoenidae.
Comments: I still haven't had a chance to read this, but this is a fantastic new contribution summarizing much of what we know, and some newly published data on fossil whales and dolphins from the Pisco Formation of Peru. Loads of incredible field photos are in this paper.

A review of the morphological patterns exhibited by all the main radiations of mysticete (baleen whale) cetaceans provided a broad assessment of the fundamental morphological transformations that occurred in the transition to the Mysticeti clade. Skull and postcranial characters were illustrated, described and compared, and their distribution was mapped on a combined phylogeny in the search for morphological support for the principal mysticete clades (i.e., Mysticeti, Chaeomysticeti and Balaenomorpha). In particular, characters of the skull (rostrum, vertex, temporal fossa, tympanic bulla and dentary) and the postcranial appendicular skeleton (scapula, humerus, radius and ulna) were all involved at different degrees in the process of morphological transformations leading to the modern-day mysticetes. Apart from a few typical characteristics of the rostrum that were already present in the earliest-diverging mysticetes (presence of lateral process of the maxilla, presence of multiple dorsal infraorbital foramina, thin lateral border of maxilla and presence of mesorostral groove), most of the other anatomical districts were unaffected by the transition so the earliest mysticetes show a number of archaeocete characters in the tympanic bulla, dentary and skull roof. The analysis of the whole dataset supported the hypothesis that the origin and evolution of mysticetes constituted a step-wise process and that the bauplan of the modern-day mysticetes was fully assembled at the level of the common ancestor of all Balaenomorpha.

A new periotic (MuMAB 240508) is described from the Tortonian of the Stirone River, Northern Italy. The new specimen is described and compared to all the known periotics of extant and extinct balaenopterid species. The new specimen shows balaenopterid characters, including the triangular anterior process, the transverse and anteroposterior elongations of the pars cochlearis, as well as in the arrangement of the endocranial foramina of the periotic. The presence of a medial promontorial groove, a lack of separation between the round window and the perilymphatic foramen together with the presence of a protruding set of medial crests suggest that this periotic may belong to an archaic and undescribed balaenopterid species. A phylogenetic analysis was performed to understand the relationships of this specimen within the broader context of balaenopterid phylogeny; this analysis showed that MuMAB 240508 belongs to a basal balaenopterid taxon that is the sister group to all the other Balaenopteridae with the exception of ‘Balaenoptera’ ryani. An ordinary least squares regression analysis showed that a relationship exists linking the length of the anterior process of the periotic and the total body length in Balaenopteridae. By means of such a relationship, the total body length of the individual to which MuMAB 240508 belonged to was reconstructed and resulted in c. 14.4 m. This individual was longer than all the other contemporaneous balaenopterid mysticetes confiming the hypothesis that the origin of large size occurred in this family earlier than previously thought.

Archaebalaenoptera eusebioi is a new late Miocene (Tortonian) balaenopterid species from the Aguada de Lomas locality in the Pisco Formation, Peru. It is dated to 8.85–7.93 Ma and is represented by a nearly complete skull with cervical vertebrae. Its inferred body length and mass are respectively c. 7 m and 7–11 tonnes. Archaebalaenoptera eusebioi is characterized by a short and wide rostrum, outward bowed dentary with typical balaenopterid characters in the posterior portion (posteriorly faced articular surface of the mandibular condyle, comparatively long distance between condyle and coronoid process, vestigial postcoronoid crest and fossa) and wide temporal fossa with scarcely protruding temporal crest and posteriorly placed posterior apex of nuchal crest. Its morphology suggests that it fed similarly to the modern balaenopterid whales but with a comparatively wider gape and, possibly, with a limited number of ventral throat grooves. Palaeobiogeographical analysis shows that Archaebalaenoptera originated in the North Atlantic and the invasion of South Pacific and Mediterranean occurred by subsequent dispersal events. Our results suggest that the North Atlantic acted as a centre of origin for many balaenopterid radiations and as a starting basin for recurrent invasions of other oceanic basins.

In 2007, a whale skeleton was found and excavated in Lower Pliocene sediments at Poggio alle Mura (southern Tuscany, central Italy). This partially complete skeleton is known by the nickname ‘Brunella’. The extracted blocks containing the fossil whale were deposited in a warehouse where they remained for nine years. A new project started in 2016 with the goal of (a) preparing and stabilising the whale bones, (b) studying its anatomy and relationships, and (c) developing educational activities to disseminate scientific information to local communities and tourists. In the years 2016–2019, the ‘Brunella’ Project gave rise to an unprecedented number of activities in terms of scientific and educational efforts. The skeleton was micro-excavated, stabilised and prepared for exhibition; primary and secondary schools as well as university students were involved in educational activities both at the preparation laboratory and in their classrooms. University students were involved in a field school on palaeontological preparation that was active for three years (1 week per year); local populations were invited to visit the laboratory during special Open days where they were able to see palaeontologists working on the project, the whale and the palaeoecosystem in which it ended its life cycle; foreign tourists visited the laboratory and were involved in guided tours; social media were extensively used to disseminate results and advertise opportunities to visit the laboratory; a national TV channel screened a documentary on the whale and the project, disseminating a wealth of scientific results to hundreds of thousands of people. This project represents a unicum in Italy and can be seen as a prototype standard of an ideal process directed at preserving an important palaeontological specimen and, at the same time, enhancing the awareness and enthusiasm of local citizens for their local geoheritage.

Vertebrate paleontology was born from the efforts of amateur and commercial fossil collectors in the nineteenth century. Amateur fossil collecting is a popular hobby in the USA, though owing to different ownership laws, American vertebrate paleontologists have less positive attitudes toward amateur collectors than in Europe where amateur and professional collectors work synergistically. Collections-based and literature surveys were conducted to evaluate the scientific contribution of amateurs to vertebrate paleontology near Santa Cruz, California. The first was a survey of museum collections identifying collector status (amateur or professional) of fossils from two formations (Santa Margarita Sandstone, Purisima Formation). The second was a comprehensive literature survey for these two stratigraphic units, documenting whether fossils were collected by amateurs or professionals. The third was a literature survey of all published (2009-2021) Cenozoic marine vertebrate records for the Pacific coast of North America (Alaska to Baja California, Mexico). The first survey indicates that amateurs have contributed the most (75.9%) to Santa Margarita Formation specimens and over a third (38.1%) of Purisima Formation specimens. These contributions are of high quality as they are included in 40% of all publications through time in the study area, and amateur-collected fossils are reported in half (49%) of all publications from the Pacific coast over the past decade. These findings indicate that amateur collectors are not only capable of collecting scientifically significant specimens, but appear to be integral to the study of Cenozoic marine vertebrates on the Pacific coast. Dismissive attitudes towards amateur collectors are clearly unwarranted. Advice for cultivating strong professional-amateur relations is provided.
Comments: I started this paper in 2016 when I was invited by Bruce MacFadden via "The Fossil Project" to present at a symposium at the Southeastern Geological Society of America regional meeting on my interactions with amateur collectors. I realized that at two localities I started collecting at years before I began my undergraduate studies in paleontology were shared by amateur and professional paleontologists - so I could make direct comparisons between the contributions from each at two museums. This study does not focus on marine mammal fossils, but does focus on some important marine mammal fossil sites and assemblages. In Santa Cruz County, many critical marine mammal fossils - including holotypes - were discovered (and even prepared) by amateur collectors. If it wasn't already obvious, I'm very pro-amateur, and if anything, work with them even more closely now that I'm in 'hog heaven' for amateur paleontology - the ridiculously fossiliferous Charleston region of South Carolina.

Boessenecker 2022. Oligocene-Miocene marine mammals from Belgrade Quarry, North Carolina.
A diverse but fragmentary assemblage of fossil cetaceans is reported from the Oligocene-Miocene Belgrade Formation of North Carolina. This assemblage preserves many odontocetes including four xenorophids (Albertocetus, Echovenator sp., cf. Cotylocara, and Xenorophus sp.), a possible waipatiid (cf. Waipatiidae), a giant agorophiid-grade dolphin (Ankylorhiza), a shark-toothed dolphin (cf. Squalodon), longirostrine “swordfish” dolphins (Eurhinodelphinidae), a longirostrine eoplatanistid dolphin (cf. Eoplatanista), a longirostrine squalodelphinid dolphin, a possible early delphinidan (Kentriodontidae), as well as an eomysticetid baleen whale (Eomysticetus sp.) and sirenian fragments. Most of these taxa are characteristic of or unique to Oligocene deposits (Xenorophidae, cf. Waipatiidae, Ankylorhiza, Eomysticetus) whereas others are more typical of early or middle Miocene deposits (cf. Eoplatanista, Eurhinodelphinidae, cf. Squalodon, Squalodelphinidae, Kentriodontidae). The Belgrade Formation at Belgrade Quarry is dated to 25.95–21.12 Ma, approximating the Oligocene-Miocene transition. The transitional composition of the Belgrade cetacean assemblage suggests gradual changes between Oligocene to Miocene cetacean faunas, to be verified by the discovery of more complete remains from the poorly sampled earliest Miocene (Aquitanian).
Comments: This study was only possible thanks to the generosity of a bunch of amateur fossil collectors, chiefly members of the North Carolina Fossil Club. This important but meager little fauna is critical as it dates to 25-21 Ma and contains surprisingly young records of Oligocene dolphins and surprisingly old records of Miocene dolphins - whether it indicates a gradual Oligo-Miocene transition in cetacean assemblages, or reworking, is unclear - though the Miocene dolphins would be anomalously old, and instead support the prior hypothesis.

A basal member of Balaenomorpha (Cetacea, Mysticeti), Persufflatius renefraaijeni, n. gen., n. sp., is described based on cranial material discovered in upper Miocene deposits of Liessel (the Netherlands). Thanks to the palynological analysis of an associated sediment sample, the specimen is dated from the late Tortonian (Dinozone SNS M14: c. 8.2-7.6 Ma). Our phylogenetic analysis recovers the new taxon at the base of the successful crown mysticete clade leading to modern rorquals. Though the holotype is only partially preserved (it consists of the partial right side of the neurocranium), it provides new data on the cranial anatomy of these early relatives of extant rorquals, which are poorly represented in the global fossil record. Several skull parts (postglenoid process of the squamosal, base of the zygomatic process of the squamosal, the anteromedioventral portion of the squamosal bone, and the exoccipital) show unusual swelling due to pachyostosis, giving the whole lateral basicranial region an inflated aspect.

Brent et al. 2022. Evolutionary assembly and disassembly of the mammalian sternum.
Evolutionary transitions are frequently associated with novel anatomical structures, but the origins of the structures themselves are often poorly known. We use developmental, genetic, and paleontological data to demonstrate that the therian sternum was assembled from pre-existing elements. Imaging of the perinatal mouse reveals two paired sternal elements, both composed primarily of cells with lateral plate mesoderm origin. Location, articulations, and development identify them as homologs of the interclavicle and the sternal bands of synapsid outgroups. The interclavicle, not previously recognized in therians, articulates with the clavicle and differs from the sternal bands in both embryonic HOX expression and pattern of skeletal maturation. The sternal bands articulate with the ribs in two styles, most clearly differentiated by their association with sternebrae. Evolutionary trait mapping indicates that the interclavicle and sternal bands were independent elements throughout most of synapsid history. The differentiation of rib articulation styles and the subdivision of the sternal bands into sternebrae were key innovations likely associated with transitions in locomotor and respiratory mechanics. Fusion of the interclavicle and the anterior sternal bands to form a presternum anterior to the first sternebra was a historically recent innovation unique to therians. Subsequent disassembly of the radically reduced sternum of mysticete cetaceans was element specific, reflecting the constraints that conserved developmental programs exert on composite structures.

Along with the transition to the aquatic environment, cetaceans experienced profound changes in their skeletal anatomy, especially in the skull, including the posterodorsal migration of the external bony nares, the reorganization of skull bones (= telescoping) and the development of an extreme cranial asymmetry (in odontocetes). Telescoping represents an important anatomical shift in the topological organization of cranial bones and their sutural contacts; however, the impact of these changes in the connectivity pattern and integration of the skull has never been addressed. Here, we apply the novel framework provided by the Anatomical Network Analysis to quantify the organization and integration of cetacean skulls, and the impact of the telescoping process in the connectivity pattern of the skull. We built anatomical networks for 21 cetacean skulls (three stem cetaceans, three extinct and 10 extant mysticetes, and three extinct and two extant odontocetes) and estimated network parameters related to their anatomical integration, complexity, heterogeneity, and modularity. This dataset was analyzed in the context of a broader tetrapod skull sample as well (43 species of 13 taxonomic groups). The skulls of crown cetaceans (Neoceti) occupy a new tetrapod skull morphospace, with better integrated, more heterogeneous and simpler skulls in comparison to other tetrapods. Telescoping adds connections and improves the integration of those bones involved in the telescoping process (e.g., maxilla, supraoccipital) as well as other ones (e.g., vomer) not directly affected by telescoping. Other underlying evolutionary processes (such as basicranial specializations linked with hearing/breathing adaptations) could also be responsible for the changes in the connectivity and integration of palatal bones. We also find prograde telescoped skulls of mysticetes distinct from odontocetes by an increased heterogeneity and modularity, whereas retrograde telescoped skulls of odontocetes are characterized by higher complexity. In mysticetes, as expected, the supraoccipital gains importance and centrality in comparison to odontocetes, increasing the heterogeneity of the skull network. In odontocetes, an increase in the number of connections and complexity is probably linked with the dominant movement of paired bones, such as the maxilla, in retrograde telescoping. Crown mysticetes (Eubalaena, Caperea, Piscobalaena, and Balaenoptera)are distinguished by having more integrated skulls in comparison to stem mysticetes (Aetiocetus and Yamatocetus), whereas crown odontocetes (Waipatia, Notocetus, Physeter, and Tursiops) have more complex skulls than stem forms (Albertocetus). Telescoping along with feeding, hearing and echolocation specializations could have driven the evolution of the different connectivity patterns of living lineages.

A nearly complete skull of Remingtonocetus harudiensis was discovered from the Harudi Formation of the Kutch Basin, western India. Though several specimens have been collected over the last two decades by earlier workers, this skull shows variations in morphology within the species that were not known earlier. Hence, this finding is significant because it helps in expanding our knowledge of the skull morphology with the addition of characters such as the larger size of the skull, the difference in dental morphology, and the two-ridged external nasal feature and our understanding of how the molar morphology can vary within the same species. The newly excavated skull is the largest Remingtonocetus skull so far discovered and is similar to the size of Dalanistes ahmedi from Kutch, India and from Baluchistan, Pakistan. Thus, the range of the overall body size of the species will change considerably towards the higher side nullifying a major distinction between Dalanistes and Remingtonocetus. The new skull was excavated from the chocolate brown shales of the clastic facies of the Harudi Formation, in the inner ramp of a lagoonal phase. It is approximately 4 m above the nodular limestones from which the other Remingtonocetus materials were previously described. The limestone represents the carbonate facies of the Formation formed in a middle to outer ramp setting. An emended diagnosis of R. harudiensis is provided in this study. The Bartonian Harudi Formation of the Kutch Basin, thus, becomes unique in having Remingtonocetus specimens discovered from two different lithological facies from both the inner and outer ramp settings.

Reaching body lengths of 9 m, killer whales (Orcinus orca) are the top mammalian predators of the present-day oceans. These distinctive, cosmopolitan dolphins feature an extremely broad and diverse trophic spectrum, ranging from mollusks like octopuses and squids to other toothed whales and even baleen whales. Due to the lack of fossils that can be confidently assigned to Orcinus or close relatives thereof, the evolutionary origin of extant killer whales has rarely been addressed. Here, we provide an updated and thorough reappraisal of the systematics, phylogeny and feeding behavior of Orcinus citoniensis from the Pliocene of Tuscany (Italy), the only fossil species of killer whale currently known on the basis of diagnostic material. Our systematic and phylogenetic reassessment confirms that O. citoniensis is a bona fide species of Orcinus that mostly differs from the extant O. orca by virtue of a smaller body size (ca. 3.5 m). Besides Orcinus spp., the subfamily Orcininae is here recognized as a relatively early-branching clade of delphinids that also includes the Italian Pliocene species “Tursiops” osennae, Hemisyntrachelus pisanus and Hemisyntrachelus cortesii. Our morphofunctional analysis supports a mainly piscivorous diet for O. citoniensis. In particular, the degree of apical tooth wear observed on the holotype is consistent with that shown by the extant generalist type of Atlantic killer whales while clearly differing from members of the cetacean-eating specialized types. The prominence of fish in the diet of O. citoniensis is further supported by the fine and shallow microwear features on the dentine exposed at the apical portion of the tooth crown. The emergence of Orcinus as one of the highest trophic level predators of the global oceans, especially at mid and high latitudes, may have involved some process of exaptation, well into the Pleistocene, when large eurytrophic sharks that used to be common and widespread in Pliocene times either became extinct or underwent a severe reduction of their biogeographic ranges.
[No figure available, I do not yet have a pdf copy of this paper]

Coombs et al. 2022. The tempo of cetacean cranial evolution.
The evolution of cetaceans (whales and dolphins) represents one of the most extreme adaptive transitions known, from terrestrial mammals to a highly specialized aquatic radiation that includes the largest animals alive today. Many anatomical shifts in this transition involve the feeding, respiratory, and sensory structures of the cranium, which we quantified with a high-density, three-dimensional geometric morphometric analysis of 201 living and extinct cetacean species spanning the entirety of their ∼50-million-year evolutionary history. Our analyses demonstrate that cetacean suborders occupy distinct areas of cranial morphospace, with extinct, transitional taxa bridging the gap between archaeocetes (stem whales) and modern mysticetes (baleen whales) and odontocetes (toothed whales). This diversity was obtained through three key periods of rapid evolution: first, the initial evolution of archaeocetes in the early to mid-Eocene produced the highest evolutionary rates seen in cetaceans, concentrated in the maxilla, frontal, premaxilla, and nasal; second, the late Eocene divergence of the mysticetes and odontocetes drives a second peak in rates, with high rates and disparity sustained through the Oligocene; and third, the diversification of odontocetes, particularly sperm whales, in the Miocene (∼18–10 Mya) propels a final peak in the tempo of cetacean morphological evolution. Archaeocetes show the fastest evolutionary rates but the lowest disparity. Odontocetes exhibit the highest disparity, while mysticetes evolve at the slowest pace, particularly in the Neogene. Diet and echolocation have the strongest influence on cranial morphology, with habitat, size, dentition, and feeding method also significant factors impacting shape, disparity, and the pace of cetacean cranial evolution.
Comments: The mind-boggling dataset for this paper includes many critical specimens from the Mace Brown Museum of Natural History. Many of the scans were produced by Morgan Churchill and Rebecca Bennion - I've since provided some photogrammetric models to Ellen Coombs for a followup study. This paper is really, really neat and a must read for any whaleontologist!

Stem cetaceans (archaeocetes) have a substantial Eocene fossil record but are largely absent since the Eocene–Oligocene boundary. Putative Oligocene archaeocetes are mostly represented by fragmentary denticulate teeth, making systematic classification difficult. The New Zealand species Kekenodon onamata (Wharekuri Greensand, Duntroonian local Stage, Chattian) is one of the most widely mentioned Oligocene archaeocetes and is the single-named species of the family Kekenodontidae. The holotype includes heterodont teeth in addition to cranial and post-cranial elements, and partial tympanoperiotics, with potential for a strong phylogenetic signal. Hitherto, the strikingly disparate Kekenodon onamata has been excluded from published phylogenetic analyses, with its affinities largely remaining speculative. Here, a detailed anatomical account allows a redescription of Kekenodon onamata, and a comprehensive phylogenetic analysis. The phylogenetic analysis shows Kekenodon onamata as the latest-surviving archaeocete, sister-taxon to the Neoceti. The large heterodont teeth with patterns of extensive enamel macrowear suggest Kekenodon onamata utilized a raptorial feeding strategy and hunted hard-boned prey, such as other marine mammals and penguins. Combined with an estimated body size of c. 8 m, Kekenodon onamata probably occupied an apex predator ecological niche similar to the modern killer whale.
Comments: I saw the original holotype fragments of Kekenodon onamata when I was a Ph.D. student, and had man discussions with Josh about his thesis research before (and after) finishing my Ph.D. It's been a long road, but the resulting paper on Kekenodon is a critical benchmark in Oligocene 'whaleontology' and I was pleased to have the opportunity to review it as well! Now Josh has to get his followup paper on the complete kekenodontid skull from NZ finished...

The polar bear (Ursus maritimus) is the apex predator of the Arctic but its distribution throughout the Pleistocene and Holocene has not previously been reported. Although natural death specimens of this species (‘fossils’) are rare, archaeological remains are much more common. This historical compilation presents the record of known ancient polar bear remains from fossil and archaeological contexts before AD 1910. Most remains date within the Holocene and derive from human habitation sites within the modern range of the species, with extralimital specimens documented in the north Atlantic during the late Pleistocene and in the southern Bering Sea during the middle Holocene reflecting natural expansions of sea ice during known cold periods. The single largest polar bear assemblage was recovered from an archaeological site on Zhokhov Island, Russia, occupied ca. 8,250–7,800 a BP during the warmer-than-today Holocene Climatic Optimum: 5,915 polar bear bones were recovered, representing 28% of all remains identified. Polar bear fossils and archaeological remains across the Arctic are most often found in proximity to areas where polynyas (recurring areas of thin ice or open water) are known today and which likely occurred in the past, including for the oldest known fossil from Svalbard (ca. 130–115 k a BP) and the oldest known archaeological specimens from Zhokhov Island (ca. 8,000 a BP). This pattern indicates that as they do today, polar bears may have been most commonly found near polynyas throughout their known historical past because of their need for ice-edge habitats at which to hunt seals.

The pan-sirenian Bauplan is conservative, probably owing to the constraints of adaptation to an aquatic lifestyle. Gathering morphological data from extinct forms is complex, resulting in poorly resolved phylogenies for stem pan-sirenians. Extant sirenians ossify the falx cerebri and the tentorium cerebelli, membranes of the dura mater of the brain attached to the parietal bone. Nevertheless, these ossifications are not present in some pan-sirenians. The basioccipital bone has received little attention in the literature except for establishing the relative age of individuals. Here, we present new cranial elements and a detailed description of the skull of Sobrarbesiren cardieli, a stem pan-sirenian from the Lutetian of Spain represented by eight individuals; we study its intraspecific variation and palaeoecological implications and explore the evolution of the endocranial structures and the basioccipital bone in pan-sirenians. Six new phylogenetic characters are added to the latest pan-sirenian dataset, resulting in a well-resolved topology where Sobrarbesiren is recovered close to the root, in a clade with Prototherium and Eotheroides aegyptiacum. The basioccipital bone and the ossified endocranial membranes have a phylogenetic signal, and the absence of such endocranial structures represents the plesiomorphic condition for pan-sirenians and is not diagnostic for the family Protosirenidae as previously believed.

Domning 2022. What Can We Infer About the Behavior of Extinct Sirenians?
Several aspects of the behavior of fossil sirenians can be inferred from their skeletal remains. Their transition from terrestrial walking to obligate swimming is relatively well documented by their postcranial skeletons. The salinity of their aquatic habitats, as well as their diets, is determinable from stable isotopes in their tooth enamel. Deflection and width of the front parts of their skulls, respectively, reflect where in the water column they fed, and how selective they were in feeding. Specializations of tusks and other teeth also offer hints about diet, intraoral food transport, and mastication. Sizes of the infraorbital and mental foramina may reflect the importance of their prehensile and tactile vibrissae. The three Recent sirenian genera have divergently adapted in radically different ways, especially in feeding adaptations, to environmental changes of the last 10 million years. Fossils shed little light on vision, chemical senses, or touch, apart from the facial vibrissae, but future study of their ear bones could reveal much about the evolution of sirenian hearing.

Fischer et al. 2022. Ecological signal in the size and shape of marine amniote teeth.
Amniotes have been a major component of marine trophic chains from the beginning of the Triassic to present day, with hundreds of species. However, inferences of their (palaeo)ecology have mostly been qualitative, making it difficult to track how dietary niches have changed through time and across clades. Here, we tackle this issue by applying a novel geometric morphometric protocol to three-dimensional models of tooth crowns across a wide range of raptorial marine amniotes. Our results highlight the phenomenon of dental simplification and widespread convergence in marine amniotes, limiting the range of tooth crown morphologies. Importantly, we quantitatively demonstrate that tooth crown shape and size are strongly associated with diet, whereas crown surface complexity is not. The maximal range of tooth shapes in both mammals and reptiles is seen in medium-sized taxa; large crowns are simple and restricted to a fraction of the morphospace. We recognize four principal raptorial guilds within toothed marine amniotes (durophages, generalists, flesh cutters and flesh piercers). Moreover, even though all these feeding guilds have been convergently colonized over the last 200 Myr, a series of dental morphologies are unique to the Mesozoic period, probably reflecting a distinct ecosystem structure.

Skeletal remains and historical accounts indicate that grey whales (Eschrichtius robustus) existed in the North Atlantic Ocean from the Pleistocene into the seventeenth century. Fossil and sub-fossil occurrences in this basin are rare, distributed from the east coast of the United States to Iceland and Europe. Here, we report an incomplete skeleton of a Holocene grey whale from Pender County, North Carolina, USA. This specimen represents a physically immature individual and is the most complete North Atlantic grey whale specimen reported to date. It comprises 42 cranial and postcranial elements, including the cranium, parts of the rostrum, both mandibles, both scapulae, humeri, radii and ulnae, most of the vertebral column anterior to the lumbar region and numerous ribs. Its provenance near the inlet of a large estuary is consistent with previous findings from the southeastern USA and parallels the species' habitat use in Baja California breeding and calving grounds in the North Pacific Ocean. Radiocarbon dating indicates an age of 827 ± 172 years before present. Cut marks on multiple skeletal elements indicate that the animal was butchered, suggesting some level of human exploitation of the species in the southeastern USA in the twelfth century, approximately 500 years prior to its extirpation in the North Atlantic.

The transition in Mysticeti (Cetacea) from capture of individual prey using teeth to bulk filtering batches of small prey using baleen ranks among the most dramatic evolutionary transformations in mammalian history. We review phylogenetic work on the homology of mysticete feeding structures from anatomical, ontogenetic, and genomic perspectives. Six characters with key functional significance for filter-feeding behavior are mapped to cladograms based on 11 morphological datasets to reconstruct evolutionary change across the teeth-to-baleen transition. This comparative summary within a common parsimony framework reveals extensive conflicts among independent systematic efforts but also broad support for the newly named clade Kinetomenta (Aetiocetidae + Chaeomysticeti). Complementary anatomical studies using CT scans and ontogenetic series hint at commonalities between the developmental programs for teeth and baleen, lending further support for a 'transitional chimaeric feeder' scenario that best explains current evidence on the transition to filter feeding. For some extant mysticetes, the ontogenetic sequence in fetal specimens recapitulates the inferred evolutionary transformation: from teeth, to teeth and baleen, to just baleen. Phylogenetic mapping of inactivating mutations reveals mutational decay of ‘dental genes’ related to enamel formation before the emergence of crown Mysticeti, while ‘baleen genes’ that were repurposed or newly derived during the evolutionary elaboration of baleen currently are poorly characterized. Review and meta-analysis of available data suggest that the teeth-to-baleen transition in Mysticeti is one of the best characterized macroevolutionary shifts due to the diversity of data from the genome, the fossil record, comparative anatomy, and ontogeny that directly bears on this remarkable evolutionary transformation.
Comments: This is one of several review papers over the past five years to cover the teeth to baleen transition, in a seesaw or tug-of-war over the narrative: did teeth and baleen overlap, or not? This is by far and away the most exhaustive and careful review but also includes a number of critical reanalyses of different cladistic matrices. My own matrix was reanalyzed, and out of the morphology-only matrices for mysticete evolution, it fared perhaps the best. I've been told that any praise is extremely difficult to earn from John Gatesy, so I wear it as a badge of pride! Easily the most important recent paper on the evolution of feeding morphology in mysticetes.

Pachycetus paulsonii, Pachycetus wardii, and Antaecetus aithai are middle Eocene archaeocete whales found in Europe, North America, and Africa, respectively. The three are placed in the new basilosaurid subfamily Pachycetinae. Antaecetus is a new genus known from Egypt and Morocco, and the only pachycetine known from a substantial postcranial skeleton. The skull of A. aithai described here resembles that of Saghacetus osiris in size, but lacks the narrowly constricted rostrum of Saghacetus. Antaecetus is smaller than Pachycetus and its teeth are more gracile. Upper premolars differ in having two rather than three accessory cusps flanking the principal cusp. Pachycetines differ from dorudontines in having elongated posterior thoracic and lumbar vertebrae like those of Basilosaurus, but differ from basilosaurines and from dorudontines in having conspicuously pachyosteosclerotic vertebrae with dense and thickly laminated cortical bone surrounding a cancellous core. Pachycetinae are also distinctive in having transverse processes on lumbar vertebrae nearly as long anteroposteriorly as the corresponding centrum. We infer from their pachyosteosclerotic vertebrae that pachycetines were probably sirenian-like slow swimmers living in shallow coastal seas and feeding on passing fish and mobile invertebrates.

CT-scans of a cetacean pathological vertebra from the Calvert Formation of the Miocene Chesapeake Group of Maryland, show features characteristic of a shear-compression fracture with comminution and significant periosteal reaction. The etiology of the injury suggests an intense hyperflexion of vertebrae in at least the lumbar region of the axial column. The trauma was sufficiently forceful to break much of the lower two-fifths of the centrum away from the anterior end of the body of the vertebra. However, the trauma was not immediately fatal as significant fusion of fragmented elements was well underway at the time of death. Much of the lateral and ventral surfaces of the centrum are covered with a thick layer of periosteal reactive bone. This reactive periosteal bone growth could be due to spondyloarthritis, infection, or from the traumatic event itself, if the direct muscle attachments on the vertebra were avulsed. A single megatoothed shark tooth from Otodus megalodon was found with the vertebra. It is not known if the tooth came to be there serendipitously, or if it was associated because it was lost as a result of the possible originating failed predation event, or during a final successful predation or subsequent scavenging event. The fractures are severe and unlikely to have had an endogenous origin like convulsions, seizures, or spasms. Seizures can cause vertebral fractures in humans, including elderly adults with poor bone health as well as physically fit younger individuals. Seizures causing injuries of this magnitude have not been observed in cetaceans, though domoic acid toxicity from harmful algal blooms are known to cause seizures in cetaceans, and are implicated in the deaths of neonatal skim-feeding mysticetes. It is unlikely, but possible, that a large mysticete would be affected by domoic acid toxicity to the point of a spinal fracture-causing seizure. Similarly, protozoal infections are known to cause seizures in cetaceans, though physical diagnosis of this is impossible in a fossil. Partly healed bone fractures of the face from possible collisions with the seafloor have been reported from fossil mysticetes of shallower regions of this fauna, but a spinal fracture this far back in the spinal column seems unlikely to be the result of a seafloor collision. Even though the cause of the vertebral hyperflexion and resulting trauma is unknown, a plausible cause was a crushing ambush delivered by a macropredatory shark or macroraptorial physeteroid. In spite of extant cetaceans being subjected to anthropogenically-induced trauma, which include vessel-strike blunt force injuries of many different kinds, shear-compression fractures and periosteal reactions like the ones detailed here have not yet been reported in extant cetaceans. Therefore, we consider the fracture as likely due to an impact from a predator, such as Otodus megalodon, or possibly from seizures due to a harmful algal bloom and resulting domoic acid toxicity. In either scenario, the cetacean survived.

Goswami et al. 2022. Attenuated evolution of mammals through the Cenozoic.
The Cenozoic diversification of placental mammals is the archetypal adaptive radiation. Yet, discrepancies between molecular divergence estimates and the fossil record fuel ongoing debate around the timing, tempo, and drivers of this radiation. Analysis of a three-dimensional skull dataset for living and extinct placental mammals demonstrates that evolutionary rates peak early and attenuate quickly. This long-term decline in tempo is punctuated by bursts of innovation that decreased in amplitude over the past 66 million years. Social, precocial, aquatic, and herbivorous species evolve fastest, especially whales, elephants, sirenians, and extinct ungulates. Slow rates in rodents and bats indicate dissociation of taxonomic and morphological diversification. Frustratingly, highly similar ancestral shape estimates for placental mammal superorders suggest that their earliest representatives may continue to elude unequivocal identification.

The fossil record of true seals in the Western Atlantic includes representatives of two subfamilies: Monachinae and Phocinae. The discovery of five unique mandibles from either the late Miocene marine (7.0–6.0 Ma) Eastover Formation or Early Pliocene (5.2–3.4 Ma) Yorktown Formation in North Carolina, USA, present a mixture of characters that are indicative of phocids in either the subfamily Monachinae or Phocinae. These mandibles were collected from the riverbed of the Meherrin River, a tributary of the Chowan River in North Carolina (USA). Detailed morphological analyses compared these newly discovered mandibles to representatives of other pinnipeds (Phocidae, Otariidae, and Odobenidae) and to representatives from phocid subfamilies (Cystophorinae, Devinophocinae, Monachinae, and Phocinae). Based on comprehensive morphological assessment and comparison to fossil and modern taxa, two new Monachinae species are identified: Homiphoca murfreesi and Hadrokirus novotini. Three new Phocinae species were also identified: Phocanella pumila, Leptophoca lenis and a phocine mandible that cannot be assigned to a specific genus or species at this time.

Helm et al. 2022. Possible Pleistocene Pinniped Ichnofossils on South Africa’s Cape South Coast.
Exposed surfaces of cemented foreshore deposits and aeolianites on the Cape south coast of South Africa have been demonstrated to contain numerous Pleistocene vertebrate tracksites. Two ichnosites have recently been identified that appear to demonstrate traces made by seals. These would be the first seal trace fossils thus far described in the global fossil record. The sites are situated 560 m apart in the Goukamma Nature Reserve. One site exhibits apparent flipper traces and a furrow, and the other site exhibits impressions consistent with moulds of juvenile seals. In conjunction with new luminescence dating of the associated sediments, these findings suggest a seal presence on Cape south coast beaches ∼75,000 years ago in the mild glacial period of Marine Isotope Stage 5a.Comments: I have not yet had a chance to read this, but I love the idea of pinnipeds flopping around on a Pleistocene beach and leaving trace fossils. Given how these deposits rarely become cemented or exposed along bedding planes, I did not ever expect pinniped trace fossils to ever be found in beach deposits!

Molecular phylogenetic studies that have included sirenians from the genera Trichechus, Dugong, and Hydrodamalis have resolved their interrelationships but have yielded divergence age estimates that are problematically discordant. The ages of these lineage splits have profound implications for how to interpret the sirenian fossil record—including clade membership, biogeographic patterns, and correlations with Earth history events. In an effort to address these issues, here we present a total evidence phylogenetic analysis of Sirenia that includes living and fossil species and applies Bayesian tip-dating methods to estimate their interrelationships and divergence times. In addition to extant sirenians, our dataset includes 56 fossil species from 106 dated localities and numerous afrotherian outgroup taxa. Genetic, morphological, temporal, and biogeographic data are assessed simultaneously to bring all available evidence to bear on sirenian phylogeny. The resulting time-tree is then used for Bayesian geocoordinates reconstruction analysis, which models ancestral geographic areas at splits throughout the phylogeny, thereby allowing us to infer the direction and timing of dispersals. Our results suggest that Pan-Sirenia arose in North Africa during the latest Paleocene and that the Eocene evolution of stem sirenians was primarily situated in the Tethyan realm. In the late Eocene, some lineages moved into more northern European latitudes, an area that became the source region for a key trans-Atlantic dispersal towards the Caribbean and northern-adjacent west Atlantic. This event led to the phylogenetic and biogeographic founding of crown Sirenia with the Dugongidae-Trichechidae split occurring at the Eocene-Oligocene boundary (~33.9 Ma), temporally coincident with the onset of dropping global sea levels and temperatures. This region became the nexus of sirenian diversification and supported taxonomically-rich dugongid communities until the earliest Pliocene. The Dugonginae-Hydrodamalinae split occurred near Florida during the early Miocene (~21.2 Ma) and was followed by a west-bound dispersal that gave rise to the Pacific hydrodamalines. The late middle Miocene (~12.2 Ma) split of Dugong from all other dugongines also occurred near Florida and our analyses suggest that the Indo-Pacific distribution of modern dugongs is the result of a trans-Pacific dispersal. From at least the early Miocene, trichechid evolution was based entirely in South America, presumably within the Pebas Wetlands System. We infer that the eventual establishment of Amazon drainage into the South Atlantic allowed the dispersal of Trichechus out of South America no earlier than the mid-Pliocene. Our analyses provide a new temporal and biogeographic framework for understanding major events in sirenian evolution and their possible relationships to oceanographic and climatic changes. These hypotheses can be further tested with the recovery and integration of new fossil evidence.

The aetiocetids are a well-known extinct taxon of Oligocene toothed mysticetes in the North Pacific. They display a high degree of diversity contrasting with that of other tooth-bearing mysticetes such as mammalodontids, llanocetids and Coronodon spp. The disparate developmental patterns (heterochrony) and origin of the first kelp forests in the North Pacific during the Eocene–Oligocene transition might explain such diversity. Aetiocetidae have been the focus of debate regarding the origin of baleen and filter-feeding in Mysticeti. The current interpretation of aetiocetid feeding strategies is based on the presence of teeth, unfused mandibular symphysis and lateral palatal foramina. However, our understating of their feeding apparatus function is limited, leaving associated structures like the zygomatic arch unnoticed. The present study describes a new fossil of a small-sized mysticete (∼3 m long), Kaaucetus thesaurus gen. et sp. nov., which belongs to Aetiocetidae. The protruded squamosal prominence and distinctive zygomatic process present in this new fossil highlight disparity in feeding morphology amongst aetiocetids. Moreover, the zygomatic arch complex and masticatory muscle system reconstruction are crucial to interpreting the likely capability of mastication (shearing motion) in aetiocetids and other toothed mysticetes. Lastly, the record of several aetiocetid genera in Mexico extends the geographical range of the group into the subtropical region.

Eomysticetids are an extinct basal taxon of baleen whales (Mysticeti: Chaeomysticeti - ‘true whales’), which appeared during the Oligocene and reflected a transitional stage between the origin of baleen and the loss of teeth, a trait that characterizes extant baleen whales. The eomysticetids are recognized as a diverse and widely distributed lineage with a rich record in the Australasia region (Australia and New Zealand). Several features of their palaeobiology, such as ontogenetic growth and ecological feeding, have been discussed; however, traits related to their biogeography and inherent speciation (origin and extinction) have only been briefly reviewed. In this context, the present study addresses biogeographic aspects based on a new fossil from the eastern North Pacific belonging to the records of the El Cien Formation of Baja California Sur, Mexico. Our description and phylogenetic analysis classify this new specimen of eomysticetid within the genus Eomysticetus (cf. Eomysticetus sp.). In addition, we identify the biogeographic relationship between the (eastern) North Pacific and the (western) North Atlantic, which indicates a possible exchange of their Oligocene cetacean faunas. Furthermore, the presence of the Gaarlandia terrestrial arc during the Oligocene indicate vicariance events, which likely led to allopatric speciation within Eomysticetidae in the North Hemisphere. Lastly, the Oligocene cetacean fossil record from Baja California Sur, unique in Mexico, might explain several aspects of the eomysticetids palaeobiology, considering the several unnamed specimens housed in the local collections.
Comments: Always love to see new eomysticetid specimens get published!

Iliopoulos et al. 2022. The Fossil Record of Sea Cows (Mammalia: Sirenia) in Greece.
Sea cows are a group of aquatic, mainly marine mammals (Mammalia: Sirenia), being common dwellers in shallow coastal and transitional environments of the former Tethys Ocean, and which are found in the fossil record from the early Eocene till present. The review of the fossil record of this group in Greece revealed the presence of a single sirenian taxon, Metaxytherium medium, belonging to the family Dugongidae. Sirenian remains have been found to date in at least 20 localities, in upper Miocene sediments mainly of Tortonian age. Notably, 18 of the reported localities have been found all over Crete, one locality is reported for the first time herein from Gavdos Island and one more also reported for the first time herein that has been found on Cephalonia Island. The most important finding is possibly the articulated skeleton found in the locality Stylos of Apokoronas province in Western Crete.

Kiel et al. 2022. Seals, whales and the Cenozoic decline of nautiloid cephalopods.*
Nautilus and Allonautilus, last members of the once widespread nautiloid cephalopods, are today restricted to the deep central Indo-West Pacific Ocean, for reasons that remain unclear. Cephalopod evolution is generally considered as being driven by vertebrate predation; therefore, we investigated the role of whales and seals in the decline of nautiloids through the Cenozoic. Distribution data for nautiloids, pinnipeds and cetaceans through the Cenozoic were compiled and plotted on a series of paleogeographic maps. Nautiloid shell sizes were compiled and plotted against the first appearance of pinnipeds and cetaceans in key regions. From the Oligocene onward, nautiloids became extinct in areas where pinnipeds appeared. The exception is the agile nautiloid Aturia, extinct globally at the end of the Miocene. A major role of odontocetes in the demise of nautiloids is not apparent, except for a few brevirostrine Oligocene taxa from the North American Atlantic and Pacific coasts, which appeared in these areas at the same time as nautilids disappeared. The Oligocene disappearance of nautiloids (except Aturia) from the American Pacific coasts coincides with the development of oxygen minimum zones (OMZs) in this region. We hypothesize that the Cenozoic spread of pinnipeds drove nautiloids into their present-day central Indo-West Pacific refuge. Additional factors for the local extinction of nautiloids in the Oligocene include predation by short-snouted whales and the development of OMZs, preventing nautiloids from retreating into deeper water.

Kimura et al. 2022. A New Species of Baleen Whale (Isanacetus-Group) from the Early Miocene, Japan.The Isanacetus-group is one of the most enigmatic groups of cetaceans. Although their phylogeny is still controversial, many previous studies suggested that they are a paraphyletic group of baleen whales, including an ancestor of Balaenopteridae and Eschrichtiidae. A new fossil from an Isanacetus-group baleen whale has been recovered from the Minamishirado Formation (latest early Miocene, Burdigalian), Iwaki, Fukushima Prefecture, Japan. The specimen consists of a cranium, mandible, and postcranial elements, including cervical, thoracic, and lumbar vertebrae, scapula, ulna, and ribs. We describe and diagnose the specimen as a new genus and species, Jobancetus pacificus. A unique combination of morphological characters characterized the specimen, e.g. frontals forming a triangular elevated plateau at the vertex, sharp, and well-developed transverse crest on the supraorbital process of frontal, sagittal crest formed by frontals and parietals, and large squamosal fossa, which is posteriorly extended well beyond the level of the posterior surface of the occipital condyle. Phylogenetic analysis under equal weighting suggests that J. pacificus is a stem group of Pligogulae, whereas the phylogenetic analyses under implied weightings suggest that J. pacificus is a stem group of Balaenopteridae + Eschrichtiidae. The discovery of J. pacificus expands our knowledge of the enigmatic Isanacetus-group.

Kimura and Hasegawa. 2022. A New Physeteroid from the Lower Miocene of Japan.
We describe a new specimen of physeteroid from the lower Miocene (Burdigalian) of Japan. This specimen was recovered from the Toyohama Formation, Chita County, Aichi Prefecture, Japan in 1984 and includes a finely preserved cranium with detached teeth and ear bones (periotic, tympanic bulla, and malleus). Here we refer this specimen to a new genus and species of the Physeteroidea, Miophyseter chitaensis gen. et sp. nov. Our phylogenetic analysis suggests that Miophyseter is a physeteroid more closely related to the crown Physeteroidea (Physeteridae and Kogiidae) than the macroraptorial physeteroids that flourished in middle and late Miocene times. A deep and large excavation on the ventral surface of the palatine and pterygoid in Miophyster suggests an adaptation for deep dives and/or the development of robust pterygoid muscles for active biting.


Eurhinodelphinids are a family of extremely long-snouted dolphins that once was widespread across the North Atlantic realm, but so far has not been recorded from the epicontinental Paratethys Sea. Here, we describe a new specimen of Xiphiacetus cristatus, including the cranium, left ear bones and a partial mandible, from the Middle Miocene (Badenian) of Austria. Our new fossil is the first record of this species outside the North Atlantic proper, and the first unequivocal record of eurhinodelphinids from the Paratethys. Its presence suggests a local invasion via the Mediterranean, and corroborates the persistence of an active marine gateway between the Mediterranean and the Central Paratethys throughout the Middle Miocene. Measurements of the bony labyrinth reveal that X. cristatus likely employed narrow-band high-frequency echolocation, making eurhinodelphinids only the second extinct odontocete family with this trait. Marked non-orthogonality of the semicircular canals suggests that X. cristatus was unsuited to rapid head movements, and thus more likely a benthic forager than a snap feeder.

Desmostylia is an extinct clade of marine mammals with two major sub-clades, Desmostylidae and Paleoparadoxiidae, known from Oligocene to Miocene strata of the North Pacific coastline. Within Paleoparadoxiidae, three genera have been identified: Archaeoparadoxia, Paleoparadoxia, and Neoparadoxia. The latter taxon is the geochronologically youngest palaeoparadoxiid and Neoparadoxia is characterized by a comparatively larger body size, although it is known only from a few specimens within a short temporal and geographic range. Here we report the discovery of an isolated tooth, which we identify as Neoparadoxia cf. N. cecilialina, constituting only the second individual specimen of Neoparadoxia with preserved dentition yet reported. This specimen was collected near Corona, California, USA, and we attribute it to the “Topanga” Formation, extending the geographic range of this taxon in Southern California. While the exact geographic locality was not recorded when it was collected in 1913, we establish two potential localities based on associated hand-written museum label and new stratigraphic information. Although initially identified as Desmostylus hesperus, this specimen of Neoparadoxia was collected 10 years before the first named paleoparadoxiid from Japan. We expect that description of more complete desmostylian material from elsewhere in Southern California will clarify the taxonomic richness and paleoecological role of this clade in Cenozoic marine mammal assemblages.

An incomplete cranium, three cervicals (including the axis) and two likely lumbars of a monodontid cetacean are here described from lower Pliocene (ca. 5.1–4.5 Ma) marine sandstones cropping out at Arcille (Grosseto Province, Tuscany, Italy). This fossil find comes from the same locality as the holotype of Casatia thermophila, which it resembles in terms of overall size and cranial morphology, and especially, by displaying a similarly depressed portion of the dorsal surface of the premaxillae anterior to the premaxillary sac fossae and medial to the anteromedial sulci. Our new find is thus assigned to C. thermophila, and significant anatomical parts that are missing in the holotype are described in order to improve the diagnosis of this monodontid species. Some dentigerous fragments of the maxillae hint at a homodont and polydont dentition, which in turn suggests a ram prey capture method that differs from the highly derived suction method that is proper of extant monodontids. This second find of C. thermophila from the warm-water Arcille palaeoenvironment lends further support to the hypothesis that monodontids once thrived in tropical and subtropical habitats.

Kolponomos newportensis is an enigmatic Miocene mammal allied to stem Pinnipedimorpha. It has been suggested that Kolponomos fed on hard-shelled benthic marine invertebrates by using its mandible as a wedge to dislodge its prey from the sea bottom by means of strong pull and torque forces. This unique feeding style was thought to originate from a singular case of mosaic convergence in mandible biomechanics between Kolponomos and the sabretoothed cat Smilodon, which complied with similarly strong torque forces when grappling with prey. As such forces must have reflected on the cranium as well, we hypothesize that the convergence between Kolponomos and the sabretoothed cats could have affected its shape. To test this hypothesis, we looked for patterns of morphological convergence in cranial shape between Kolponomos and sabretoothed cats. We found that Kolponomos is not distinctly closer to Smilodon than a number of other pinnipeds. Yet, local areas of shape convergence with Smilodon are observed in the canine area and the posterior part of the cranium, that is where the bite applies and the temporalis muscle is located, respectively. These results indicate that the mosaic convergence present between the mandibles of Kolponomos and Smilodon is partially reflected in the cranium as well.

Comments: One of the more humble but personally one of the papers I was most excited for this year - a review of fossil marine mammals from the Monterey Formation and their biostratigraphic ages. Much of this data was collecting dust in LACM for decades, known only to LACM paleontologists until recently, when the 'vault' was finally opened up (so to speak).

Peredo et al. 2022. Lateral palatal foramina do not indicate baleen in fossil whales.
Today’s mysticetes filter-feed using baleen, a novel integumentary structure with no apparent homolog in any living mammal. The origins of filter-feeding and baleen can be informed by the fossil record, including rare instances of soft tissue preservation of baleen and also by potential osteological correlates of baleen. Lateral palatal foramina on the roof of the mouth have been proposed as potential osteological correlates of baleen and their presence in some tooth-bearing stem mysticetes has led to the hypothesis that these early mysticetes possessed both teeth and incipient baleen. Here, we test this hypothesis by examining lateral palatal foramina in both filter-feeding and non-filter-feeding cetaceans, including crown and stem odontocetes and in stem cetaceans (or archaeocetes). We also confirm the presence of lateral palatal foramina in 61 species of terrestrial artiodactyls. CT scanning demonstrates consistent internal morphology across all observed taxa, suggesting that the lateral palatal foramina observed in extant mysticetes are homologous to those of terrestrial artiodactyls. The presence of lateral palatal foramina in terrestrial artiodactyls and non-filter-feeding whales (odontocetes and archaeocetes) suggests that these structures are not unique predictors for the presence of baleen in fossil whales; instead, these structures are more probably associated with gingiva or other oral tissue.

Herein we describe a new finding of a medium-sized sperm whale from the Burdigalian (Lower Miocene) of the Pietra leccese formation (southern Italy) on the basis of a partly prepared specimen that includes a partial cranium, seven detached teeth, the fragmentary right mandible and two partial vertebral bodies. Because of the overall compression of the specimen, we carried out a retro deformation of a 3D model of the cranium obtained via CT-scanning. The combined analysis of the original specimen and the retrodeformed model has allowed us to recognise that the studied specimen constitutes a new physeteroid taxon: Angelocetus cursiensis n. gen. n. sp., a longirostrine sperm whale characterised by a sideward projected supracranial basin, as evidenced by the overall displacement of its posteriormost margin. Based on a phylogenetic analysis, A. cursiensis n. gen. n. sp. is determined to be a crown physeteroid that does not belong to either the Physeteridae or the Kogiidae. The wide temporal fossa, elongated rostrum and slender teeth, as well as the skull dimensions (estimated bizygomatic width c. 550 mm) suggest a diet based on medium to large-sized bony fish that were likely captured by a raptorial pierce feeding strategy (as for most of the coeval Burdigalian physeteroids). Despite a seemingly low ecomorphological disparity, the high degree of taxonomic diversity of the Burdigalian physeteroids suggests that this time span represents a crucial phase for the evolutionary history of sperm whales.

Peters et al. 2022. A late Miocene balaenopterid petrotympanic from Liessel (The Netherlands).
In 2020 a new fossil species of rorqual (Balaenopteridae) was described as Archaeobalaenoptera liesselensis Bisconti, Munsterman, Fraaije, Bosselaers & Post, 2020 from a sandpit in Liessel (Province of Noord-Brabant, The Netherlands) based on a skull stored in the Oertijdmuseum in Boxtel. The collection of Museum Klok & Peel in Asten (The Netherlands) holds a well preserved unidentified rorqual petrotympanic from the same sandpit. A comparative study of this important skull element showed that it probably belongs to the same species. Our comparison also proved again that during the late Tortonian at least two, maybe even more, different rorqual species were already frequenting the southern North Sea Basin.

One of the largest and least documented populations of dugongs (Dugong dugon) resides in the coastal waters of the United Arab Emirates, and waters surrounding Saudi Arabia, Bahrain, and Qatar. The archaeological record of dugongs in the Gulf Region is abundant, but little is known about their fossil record in the region. Here we report an isolated sirenian rib fragment from the Futaisi Member of the Fuwayrit Formation near the town of Al Ruwais, in northern Qatar. The Fuwayrit Formation is a marine Pleistocene deposit exposed onshore in Qatar and the United Arab Emirates. Based on the correlative dating of the basal Futaisi Member with other onshore platforms, the rib fragment is approximately 125 ka. We propose that this isolated rib (likely the first rib from the right side) belongs to Dugongidae, with strong similarities to extant Dugong. We cannot, however, eliminate the possibility that it belongs to an extinct taxon, especially given its similarities with other fossil dugongid material from both Qatar and elsewhere in the world. Aside from reflecting the presence of Gulf seagrass communities in the Pleistocene, this occurrence also suggests that different (and potentially multiple) lineages of sirenians inhabited the Gulf Region in the geologic past.

Xenarthra are, very probably, the most basally branching clade among Placentalia, and the result of the so-called “Splendid Isolation” of South America. Although this is the only endemic clade with living representatives, its current diversity is very poor compared with that achieved during most of the Cenozoic. One particular clade, Folivora (living and fossil sloths), includes one of the most enigmatic animals, Thalassocnus, the only genus of the subfamily Thalassocninae. It was originally interpreted as an aquatic sloth taking into account the previous records, which are limited to coastal areas of the latest Miocene and Pliocene of Peru and Chile. Here we present the first record of Thalassocnus from Argentina, which in turn represents the first finding of the genus in a continental sequence. The materials (JUY-P-0180, radius, ulna and part of the articulated left manus) were exhumed from the upper member of the Tafna Formation (Late Miocene–Pliocene) in the eastern Puna (Jujuy Province), Argentina. In agreement with our comparative study, the cladistic analysis corroborates its identification as belonging to the genus Thalassocnus, being placed in a relatively basal location. This finding is of paleobiogeographic and paleoenvironmental significance, due to its location, more than 1171 km away from the Peruvian locality (Sacaco) and more than 750 km from the nearest ones in Chile (Caldera). This extends the geographic range from 76°W to 65°53′W, and from typical littoral environments, to clearly more terrestrial ones. The evolutionary and paleobiogeographic history of Thalassocnus is more complex than previously interpreted.

The fossil record of true seals (Family Phocidae) is notoriously poorly preserved, most notably in the Southern Hemisphere. This fossil record bias has made it difficult to assess whether populations of phocid species in the Southern Hemisphere remained resident in particular regions throughout deep time, with only the eastern South Pacific preserving a near-continuous fossil record. Here we report two new seal fossil remains from the late Miocene-early Pliocene of southeastern Australia, discovered from a new site, Black Rock. This is the fourth phocid fossil site in Australia, and the seventh in Australasia. The two fossils are referrable to the clades Pinnipedia and Monachinae. Combined with other recent fossil phocid finds in Australasia, this suggests that true seals were continuously present in the region at least throughout the Pliocene. This lends further support for the importance of Australasia in the global evolution of true seals.

Rule et al. 2022. True seals achieved global distribution by breaking Bergmann’s rule.
True seals (phocids) have achieved a global distribution by crossing the equator multiple times in their evolutionary history. This is remarkable, as warm tropical waters are regarded as a barrier to marine mammal dispersal and—following Bergmann's rule—may have limited crossings to small-bodied species only. Here, we show that ancestral phocids were medium sized and did not obviously follow Bergmann's rule. Instead, they ranged across a broad spectrum of environmental temperatures, without undergoing shifts in temperature- or size-related evolutionary rates following dispersals across the equator. We conclude that the tropics have not constrained phocid biogeography.
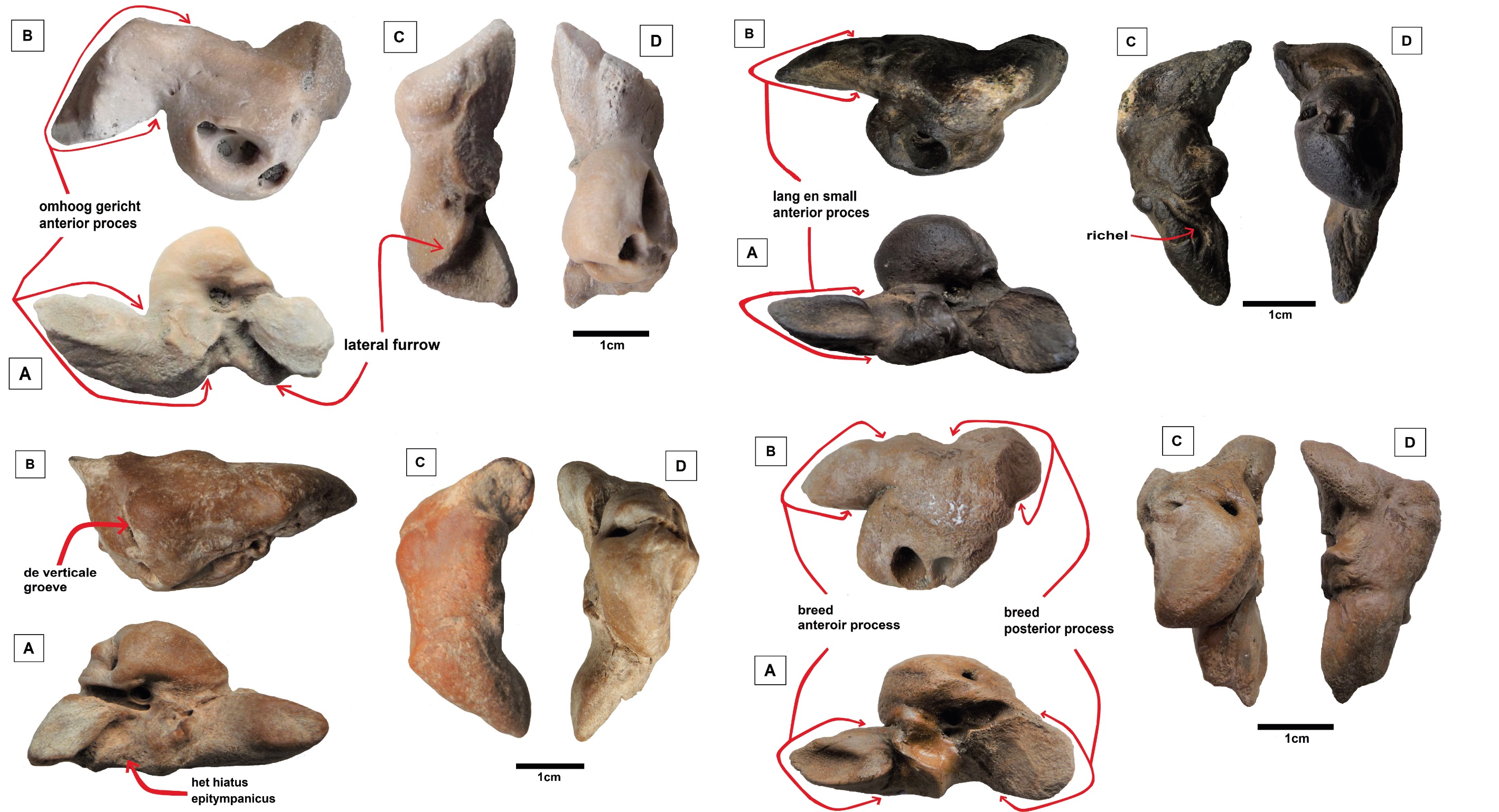
This article describes the fossil periotic (or petrosal) bones of toothed whales (Odonticeti) that are known and can possibly be found in the Dutch fossil record. The toothed whale families that are being discussed in this article are the sperm whales (Physeteridae), pygmy sperm whales (Kogiidae), Indian river dolphins (Pomatodelphininae), shark-toothed dolphins (Squalodontidae), long-snouted dolphins (Eurhinodelphinidae), beaked whales (Ziphiidae), archaic dolphins (Kentriodontidae), Franciscan dolphins (Pontoporiidae), oceanic dolphins (Delphinidae), porpoises (Phocoenidae) and the beluga whales and narwhals (Monodontidae). This article also sums the diagnostic characteristics of the toothed whale periotics and discusses the challenges they bring when studying these fossils.
Comments: Jesus I really wish I could read Dutch. My next blog post is similar to this paper... I think!

Baleen whales have lost their functional teeth and begun to use their baleen plates to feed on small prey. Modern baleen whales exhibit different types of feeding strategies, such as lunging, skimming and so on. The evolution of feeding strategy in the Chaeomysticeti is an important step in considering niche partitioning and diversification, feeding efficiency and gigantism, and evolution and extinction. This study analyses the rostrum morphology to test the hypothesis that specific rostral morphologies facilitate special feeding strategies, using modern species and their observed feeding strategies. By this means, the convergence of rostral morphology can be recognized in the closest groups in the morphospace. As a result, the two linages (Balaenidae and Caperea marginata) are recognized to have convergent rostral morphology. In addition, an early member of the Chaeomysticeti, Yamatocetus canaliculatus, and most fossil species are plotted in or close to the cluster of lunge feeders. The original feeding strategy of the Chaeomysticeti could be more similar to lunge feeding than to skim feeding. Fossil relatives of the two linages showing transitional conditions indicate that they shifted to skim feeding independently. The evolution of the feeding strategy of the Chaeomysticeti is possibly more complex than that was thought.
[no figure available, I don't have a pdf yet]

The major increase in cetacean brain size happened in the middle Miocene, about 15 million years ago, and involved the modern oceanic dolphin lineage Delphinoidea. In this paper, we describe a new specimen of an archaic dolphin, aff. Prosqualodon davidis, from the Gee Greensand, near Oamaru, New Zealand. The specimen is from the early Miocene, approximately 23–19–18 Ma, and includes an incomplete cranium, endocast, teeth, and some postcranial elements. Comparison of the median sulcus among previously reported endocasts and brains of the Cetacea suggests that the cerebrum was expanded incipiently in the Odontoceti from the early Miocene in the Eurhinodelphinidae + Delphinoidea lineage. Conversely, the olfactory fossa has been reduced in Odontoceti, and completely lost in at least two clades (Delphinoidea and Platanista gangetica), because olfaction was most likely no longer an essential sense for animals spending their life in water. Reduction and loss of the olfactory fossa saved energy, which may be related to the expansion of the cerebrum and cortex in an Odontoceti lineage that led to modern oceanic dolphins. The early Miocene appears to have been a transitional period for archaic and modern-type odontocetes, which were possibly and partly separated by these differences.

New tarsals (astragalus and entocuneiform) of fossil pinnipeds from the lower Miocene (Burdigalian) Oi Formation in Mie Prefecture, Japan, belonging to early pinnipedimorphs (i.e., enaliarctines or basal odobenids) and cf. Odobenidae gen. et sp. indet. were discovered. A comparison of early pinnipedimorph astragali conclusively indicated that at least three species were represented in the western North Pacific during the early Miocene (Burdigalian). Early pinnipedimorphs clearly showed considerable diversification in the eastern North Pacific during the early Miocene, and our study demonstrates that the early pinnipedimorphs in the western North Pacific also started to diversify around the same time.

The Caucasus is one of the most important areas for the study of Paratethyan Neogene phocids, with a number of ‘paleorookery’ sites that are still poorly studied. In this study, the remains of a fossil phocine from the Late Miocene locality of Eldari I (late Vallesian/late Sarmatian s.l.) in the Southern Caucasus are described and compared with other Paratethyan phocines. The new materials obtained from the site consists of the right complete forelimb bones along with the carpal and metacarpal bones in natural anatomical articulation and a right innominate bone fragment. Additionally, published remains of ‘Phoca’ procaspica from Eldari I are analysed. Based on morphological and morphometric studies, the Eldari I phocine is assigned to the genus Praepusa. The skeletal material demonstrates a distinct combination of diagnostic traits that can be attributed to the smallest representative of the subfamily, Praepusa procaspica. During the early Late Miocene (10 Ma.), major palaeoecological and palaeogeographic changes took place in the Eastern Paratethyan basin, which significantly decreased the diversity of phocids and had a negative impact on the remnant taxa; this was expressed in the Eldari I and Küçükçekmece seals by the decrease in body size and a change in the feeding apparatus.

Van Vliet et al. 2022. Eocene cetaceans from the Belgian-Dutch coastal waters.
Thirteen cetacean vertebrae and a vertebral neural arch fragment are described and figured. All have been dredged from the southern part of the North Sea, in the tidal channels Het Scheur, Belgium, and Wielingen in the estuary of the Westerschelde, near the Belgian-Dutch border. Here, strata of the Middle Eocene Maldegem Formation and of the Late Eocene parts of the Zelzate Formation crop out at the seafloor. The vertebral centra are mostly large and show basilosaurid characteristics. Based upon morphology, three different main types are recognised. The first morphotype apparently represents a new taxon, the second one closely resembles a vertebra of a cetacean from the Priabonian of Maastricht, the Netherlands and the third is assigned to the genus Pachycetus which taxon is, amongst others, also known from the Bartonian and maybe Priabonian of Ukraine and Germany in Europe. In the vertebrae of the latter morphotype, maybe two subtypes, a large and a smaller one can be discerned. With at least three different large taxa, the seafloor at Wielingen and Het Scheur appears to be a surprisingly important site of Eocene cetacean remains in Europe.

Platanistoidea remains one of the most evolutionarily intriguing lineages of toothed whales (Odontoceti). The clade comprises mostly extinct species from the late Oligocene–early Miocene onward and a single extant riverine genus (Platanista). There is an ongoing debate as to the membership of Platanistoidea and the causes of their near extinction. In Patagonia (Argentina), the most abundant platanistoid recorded in the lower Miocene Gaiman Formation is Notocetus vanbenedeni, first described by Moreno in 1892 based on two individuals. The goal of the present contribution is to conduct an updated anatomical, palaeobiological and phylogenetic analyses of Notocetus vanbenedeni and hence contribute to an understanding of the evolutionary history of the Platanistoidea. Our analyses, including at least 26 individuals (12 undescribed), show that Notocetus vanbenedeni is a valid platanistoid taxon, recovered as part of a new clade. Among its most outstanding features, this taxon has an elevated dorsal tubercular supraorbital crest formed mainly by the frontal, the precursor of the pneumatized crest of the extant Platanista. Notocetus vanbenedeni also shows initial stages of the plesiomorphic bony connection between the earbones and skull as in Platanista, although the functional implications for hearing remain elusive. The nasal sac system, pterygoid sinus system and morphology of the earbones suggest that this species was able to hear high-frequency sounds and echolocate underwater, similar to extant odontocetes. Thus, Notocetus vanbenedeni presents a mosaic of features that suggest an intermediate platanistoid morphotype. Anatomical differences and phylogenetic analyses suggest that Peruvian specimens could not be referred to this species. The feeding apparatus of Notocetus vanbenedeni makes it the only combination suction-feeder recorded in the early Miocene of Patagonia and among the smallest odontocetes. Finally, the abundant records of Notocetus vanbenedeni in an inner shelf environment with freshwater influence suggest a possible early preference for such protected habitats.
Comments: Spectacular specimens and an amazing sample size; I just wish there were more photos of these specimens! Too few papers on fossil marine mammals focus on singletons; this is an important study that seeks to move beyond isolated specimens.

Weems et al. - Old Church Fm. fossil cetaceans
Remains of six types of Oligocene cetaceans have been recovered from the outcrop region of the Old Church Formation in the Atlantic Coastal Plain of east-central Virginia. These remains provide the first glimpse of the cetacean fauna that lived north of modern Cape Hatteras during the Oligocene. Two kinds of mysticetes are represented (aff. Coronodon sp. and Micromysticetus sp.), and four kinds of odontocetes (cf. Ankylorhiza sp., cf. Xenorophus sloanii, Xenorophidae aff. Cotylocara and Echovenator, and aff. Eosqualodon sp.). In detail, the mysticete portion of this fauna is different and somewhat less advanced than the mysticete fauna found in the Givhans Ferry Member of the Ashley Formation, indicating that the Old Church Formation probably is correlative with either the middle or lower member of the Rupelian Ashley Formation in South Carolina. The odontocete portion of this fauna is also compatible with the middle or lower members of the Ashley Formation, except for Eosqualodon sp. which probably came from a Chattianage “phantom unit” that is correlative with the Chandler Bridge Formation of South Carolina but now eroded away.
Comments: This is one of the last outstanding projects that Al Sanders' was attached to, and I was humbled to be invited by Rob Weems to help finish the work. Some of the identifications need revision, but it's done; owing to the pandemic, I was unable to go visit collections at CMM and VMNH to examine these specimens.

An almost complete and partially articulated skeleton of an Early Pleistocene baleen whale is here described. The fossil, measuring 11 m in length, was discovered in the Calcarenite di Gravina Formation at Lama Lamasinata site (Bari, southern Italy) in 1968. The bifurcated first rib combined with other characters supports the identification of the fossil whale as a possible undescribed species of Balaenoptera (Mysticeti, Balaenopteridae), close to or nested in the B. borealis–B. edeni–B. ricei clade. However, the limited number of preserved diagnostic characters suggests a prudent assignment of the Bari whale to Balaenoptera sp. The associated molluscs suggest a mid-shelf setting deposition near to the boundary between infralittoral and circalittoral environments, probably 40–60 m deep. An associated Carcharodon carcharias tooth (the first case of a possible trophic interaction between white shark and cetaceans in the Pleistocene) and shark bite marks on a rib support the hypothesis that an early scavenger action prevented the rising of the whale carcass because of the removal of abdominal tissues and the consequent reduction of the decomposition gas accumulation. The occurrence of chemosymbiotic bivalves near the skeleton could testify the development of the sulphophilic stage during decay. Overall, the Bari whale skeleton and its associated fossil fauna represent the first well-documented case of Pleistocene whale fall community. The Bari specimen sheds new light on the diversity and disparity of the mysticete fauna in the Mediterranean Pleistocene also related to the geodynamic, palaeoclimatic and palaeoceanographic conditions that favoured upwelling events and nutrients supply into the southern Adriatic basin.

Two sirenian species are present in the late Eocene Samlat
Formation near Ad-Dakhla in southwestern Morocco.
A well preserved mandible with left and right dentaries belongs to a new
protosirenid genus and species Dakhlasiren marocensis closely related to the
genus Protosiren. An early dugongid of uncertain identification (cf.
Eotheroides sp.) is also present, represented by vertebrae and ribs.
Protosirenids differ from dugongids in the form of the brain, size and
separation of nasal bones, and conformation of the anterior mandible.
Protosirenids also differ in having vertebrae with larger neural canals, in
having ligamentous rib articulations, and in lacking the pachyostotic ribs
characteristic of dugongids. We tentatively interpret the latter differences to
be related to feeding on softer vegetation farther offshore, with a thoracic
rete mirabile for counter-current heat exchange and a collapsible rib cage to
enable deeper dives. Dakhlasiren seemingly carried the divergent
specializations of Protosiren a step farther by reduction of tongue musculature
and loss of a masticatory surface at the front of the mandible.