In early spring 2011, just as I was
finishing up my master’s degree at Montana
State, I received an email from Tom
Deméré, the paleontology curator of the San Diego Natural History Museum,
inviting Morgan and I to study a new fossil of Pelagiarctos from the
“Topanga” Formation. Fortunately, I would get a chance to examine it closely in
person soon afterward – in June, I would be attending and presenting my
master’s taphonomy research at the 6th triennial conference on
secondary adaptations of tetrapods to life in the water (usually abbreviated
SATLW or simply referred to as the aquatic tetrapods conference), which was
being hosted by Tom Deméré and Annalisa Berta at San Diego State University and
the museum.
Although I had successfully
delivered my master’s defense presentation and graduated without a hitch a
month and a half prior, I was still nervous to give my presentation because it
was in front of a totally different audience – technically, the conference was
about secondary adaptations, and I was giving a talk on taphonomy. However, I
tooled it towards what we can reasonably infer from the marine vertebrate
fossil record, including about exactly how aquatic organisms were based on
their preservation – which, I concluded at the time was not much. The talk was
also fairly long; although I had 18 minutes to speak, which is fairly long, I
had not had the time to shorten it. 36 hours before driving down I-5, I was on
the beach at Bolinas prospecting with Dick Hilton when I got a funny phone
message from Tom ‘asking’ me if it would be okay to move my talk to the first
day; so I said my goodbyes to Dick and raced home down Highway 1 so I could
spend a day and a half polishing the presentation off. And then worried half
the drive down I-5 that I didn’t shorten it enough.

A comparison of the new Topanga Formation specimen (A) and the holotype (B) of Pelagiarctos.
The talk went without a hitch, and
later in the conference Morgan and I were able to sequester a few hours in the
SDNHM type room to examine the new specimen of Pelagiarctos. It
consisted of a fragmentary pair of mandibles, with the left mandible being
nearly complete and having much of its dentition (missing only a premolar, the
two molars, and an incisor). Unlike the type specimen from Sharktooth Hill
(which Morgan and I got a chance to examine in person at LACM in January 2012),
these mandibles were not fused together at the symphysis (intermandibular
joint). Symphyseal fusion is not common in modern pinnipeds, where it is
restricted to the modern walrus. I’ve also seen pathologic (diseased) mandibles
of modern otariids where, due to some bone disease, the symphysis has fused
along with a large mass of bone at the chin, accompanied by incisor and canine
loss.

The left mandible of the Topanga Formation specimen of Pelagiarctos. From Boessenecker and Churchill (2013).
The teeth present in the new
specimen confirm that the large teeth referred to Pelagiarctos thomasi
by Barnes (1988) were correctly referred. It’s not so surprising, since you
could predict the mandible shape from the teeth: they are like giant versions
of Neotherium teeth, and the mandible is like a giant Neotherium
jaw. I never really doubted Barnes’ identifications – but it was nice to confirm
them. The mandible of this new specimen is damned huge – it’s wide, deep, with
a short toothrow. The canines are robust, again with grooves on the lateral and
medial surfaces, giving the canine a figure-eight shape in cross section. The
premolars are large, primitively retaining what’s called the metaconid cusp;
most modern pinnipeds have teeth that are unicuspid (single cusp, usually
conical), with small anterior and posterior accessory cusps in otariids. The
main cusp (on lower postcanine teeth) is called the protoconid. The anterior
and posterior cusps are the remnants of the paraconid (anterior) and hypoconid
(posterior) cusps. The metaconid cusp is a fourth cusp which is present in many
primitive pinnipeds, such as the early enaliarctines, as well as Proneotherium,
Neotherium, and Pelagiarctos. The metaconid is located just
behind the principal cusp (protoconid). Many modern phocids primitively retain
all four cusps – the harbor seal is an excellent example. The crabeater seal
additionally bears a number of extra “neomorphic” (=new or novel structure)
cusps on the posterior tooth crowns, which are used for filter feeding.

The dentition of the Topanga Formation specimen. From Boessenecker and Churchill (2013).

More details of the dentition of the Topanga Formation Pelagiarctos, from Boessenecker and Churchill (2013); I had a fun time drawing the medial view of those teeth.
In addition to having these
primitive features, a couple of new features not seen in earlier walruses are
present: a lingual cingulum with small little “crenulations” forming a sawtooth
type pattern, and the presence of a labial cingulum. A cingulum is a thickened
ridge of enamel at the base of a tooth crown. Pelagiarctos is the only
walrus with a labial (cheek side of the tooth) cingulum, and only one other
walrus has a crenulated lingual (tongue side of the tooth) cingulum – the late
Miocene walrus Imagotaria downsi. At this point the uninitiated reader
might ask ‘what exactly makes this thing a walrus?’ The truth is, for the earliest
known walruses, the only synapomorphies allowing identification as a member of
the Odobenidae (walrus family) are skull features. Many of the features of the
known specimens of Pelagiarctos appear in some sea lions – such as a
mandible that is deepest near the canines. Although the fossils don’t have any
specific features that are diagnostic at the family level – several features of
the dentition are only found in early diverging “imagotariine” walruses. The
Imagotariinae was a subfamily named by Ed Mitchell and used extensively in
various papers by Barnes, but as pointed out by several studies over the past
two decades it is a paraphyletic assemblage of primitive walruses.
Nevertheless, it is a useful vernacular term; imagotariines are sea lion-like with
primitive dentitions, and ranged in size from harbor seal size (Proneotherium)
to elephant seal size (Pontolis magnus).

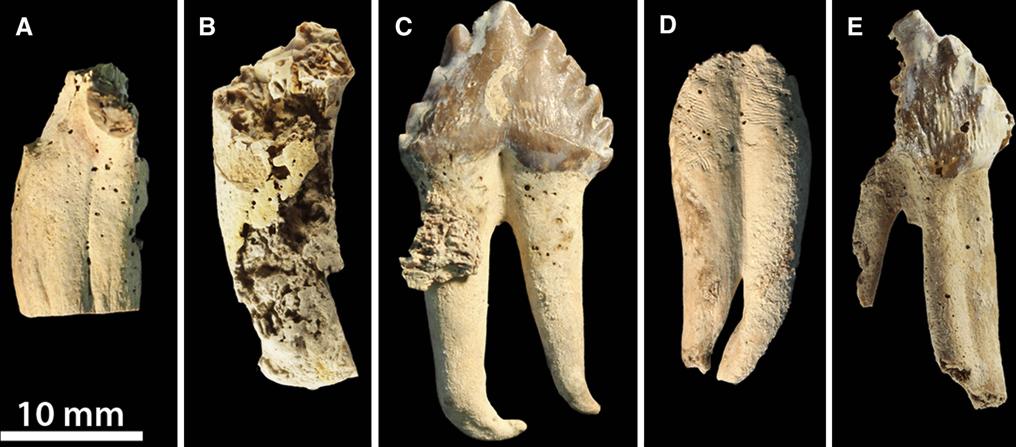
Comparison of walrus mandibles, including the Topanga Fm. Pelagiarctos specimen, Imagotaria downsi from the late Miocene Santa Margarita Sandstone of Santa Cruz County, Proneotherium repenningi from the early middle Miocene Astoria Formation of Lincoln County, Oregon, and Pontolis magnus from the late Miocene Empire Formation of Coos County, Oregon. From Boessenecker and Churchill (2013).
Because our new specimen was more
complete than the holotype, we were able to include Pelagiarctos within
a phylogenetic analysis for the first time. Previous analyses did not use many
mandibular characters, so at first we constructed a matrix which focused on
mandibular and dental characters, and only used pinniped species known by lower
jaws (i.e. we didn’t include some species for which jaws were unknown). This
meant we didn’t initially include the early walruses Prototaria and Pseudotaria
from Japan.
We originally did this because we felt we’d get more accurate results than if
we included Pelagiarctos in an analysis where it couldn’t be coded for
any cranial characters – it was a reasonable hunch at first. One of our
reviewers suggested we use a more comprehensive dataset, so we merged our data
set with that of Naoki Kohno’s (2006) analysis for his Pseudotaria muramotoi
paper. We ended up with fantastic results, and better support for some of the
relationships.

Cladograms from Deméré (1994), Kohno (2006), and our new study showing the varying position of Pontolis (underlined in red).
Most of the relationships in our
analysis are consistent with previous studies like Deméré (1994), Deméré and
Berta (2001), and Kohno (2006), with one exception. In Deméré (1994), Pontolis
magnus grouped as a dusignathine walrus, and closely related to Dusignathus
itself. In Kohno (2006), Pontolis instead formed a sister taxon
relationship with Imagotaria. The Imagotaria-Pontolis clade is only one
node below the dusignathines, so admittedly it is not a far distance. In our
analysis, however, Pelagiarctos formed a sister taxon relationship with Imagotaria
instead, based on two features: grooved canines, and a crenulated cingulum.
Neither of these features are present in Pontolis. Instead, Pontolis
plotted out as the last diverging “imagotariine” and the sister taxon to the
dusignathine + odobenine clade – in other words, intermediate between the
phylogenetic hypothesis of Deméré (1994) and Kohno (2006). It’s sort of a
compromise between the two, in a way. Obviously, there are more cranial
characters that need to be explored and new walruses to describe, so there is
clearly further scope for a more comprehensive study of walrus phylogenetics,
which is in the early planning stages.
Next up: the dramatic conclusion to
this series on the new publication, focusing on the feeding ecology of Pelagiarctos, and the life restoration.
References:
Barnes LG (1988) A new fossil pinniped (Mammalia: Otariidae) from the middle
Miocene Sharktooth Hill Bonebed, California. Contributions in Science,
Natural History Museum of Los Angeles County 396: 1–11.
Miocene Sharktooth Hill Bonebed, California. Contributions in Science,
Natural History Museum of Los Angeles County 396: 1–11.
Boessenecker, R.W. and M. Churchill. 2013. A reevaluation of the morphology, paleoecology, and phylogenetic relationships of the enigmatic walrus Pelagiarctos. PLoS One 8(1) e54311. doi:10.1371/journal.pone.0054311.
Deméré TA (1994) The family Odobenidae: a phylogenetic analysis of living and
fossil forms. In: Berta A, Deméré TA, editors. Contributions in Marine Mammal
Paleontology honoring Frank C Whitmore, Jr: Proceedings of the San Diego
Society of Natural History. 99–123.
fossil forms. In: Berta A, Deméré TA, editors. Contributions in Marine Mammal
Paleontology honoring Frank C Whitmore, Jr: Proceedings of the San Diego
Society of Natural History. 99–123.
Kohno N (2006) A new Miocene odobenid (Mammalia: Carnivora) from
Hokkaido, Japan, and its implications for odobenid phylogeny. Journal of
Vertebrate Paleontology 26: 411–421.
Hokkaido, Japan, and its implications for odobenid phylogeny. Journal of
Vertebrate Paleontology 26: 411–421.