
Happy Holidays, Merry Christmas, Happy Hannukah, Kwanzaa,
etc. – and happy Boxing Day from New Zealand.
As an American, I’m not really sure what I’m supposed to do today, so other
than recoup from consuming a disgusting amount of chicken, potatoes, cobbler,
and berry pie, I’m completing my longest ever blog post. I’ve summarized every
paper in marine mammal paleontology that has come out in 2013 (either published
or appeared online as an in press manuscript). I did this last year for 2012,
and it quickly became apparent that I left quite a few out. So, I’ve put this
rather large body of text and images together, and I’m quite frankly a bit
embarrassed with how much time I put into it, but to be fair – there were quite
a few studies that came out this year, and it forced me to read or skim several
that I had not yet had a chance to flick through. There are also several new
papers I’ve put out this year which have also inflated the list a little bit
(and there is one more yet to come this year). I hadn’t originally considered
to include my own work, but my wife convinced me to at the last minute.

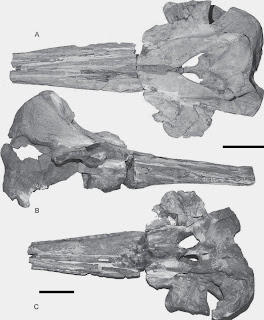
Skull of the newly described phocid Hadrokirus from the Mio-Pliocene of Peru.
Amson, E. and C. de Muizon 2013. A new durophagous phocid (Mammalia: Carnivora) from the late Neogene of Peru and considerations on monachine seals phylogeny. Journal of Systematic Palaeontology (online early).
This year’s
only new species of fossil pinniped is a new species of monachine true seal
from the latest Miocene Pisco Formation of Peru (Sud-Sacaco level). The
material is pretty well preserved, and includes several complete skulls and
mandibles, in addition to postcrania (much of which remains to be figured). Hadrokirus
– meaning stout tooth – shares a sister-taxon relationship with another Pisco
Formation phocid, Piscophoca pacifica.
The cheek teeth of Hadrokirus are very robust, and owing to this – and
some robust muscle attachments on the cervical vertebrae – Hadrokirus
was interpreted as having durophagous diet, potentially feeding on hard shelled
prey like crustaceans, or even other vertebrates. I’ll admit I was initially a
bit skeptical of the feeding ecology hypothesis, since dental features were the
rationale behind the “killer walrus” macrophagous apex predator hypothesis
initially proposed for Pelagiarctos, and criticized in the PLOS One
article published by Morgan Churchill and myself early this year. However, in
the case of Hadrokirus, there are some peculiarities of the neck muscle
attachments that add some credence to the hypothesis. Another interesting
aspect of the study is that it recovered Acrophoca as sister to extant
lobodontines (Antarctic phocids), and Hadrokirus, Piscophoca, and
Homiphoca (South Africa)
as a clade sister to the lobodontines. If this group gets further support in
the future, we could very well see a “Piscophocinae” or “Homiphocinae”.


Subfossil bowhead whale remains from Sweden.
This study reports a latest Pleistocene bowhead whale
skeleton from marine sediments along the coast of Sweden.
The skeleton is approximately 13,800 years old based on radiocarbon dating, and
includes mandibles, vertebrae, and ribs. Ancient DNA
was recovered from the specimen, which indicated that the skeleton was in fact
from a bowhead whale, Balaena mysticetus. Previously reported subfossil
bowhead remains had been used to erect the fossil species Balaena
swedenborgii; morphologically, the specimen did not appear to represent Balaena
swedenborgii, but this fossil species has been interpreted by some to
perhaps be a synonym of Balaena mysticetus or perhaps a subspecies of it
(or alternatively Eubalaena glacialis). The new fossil is probably not
complete enough to weigh in on the taxonomic distinctiveness of B.
swedenborgii anyway. The authors also inspected the specimen for any
evidence of colonization by the bone-eating worm Osedax – and found
none. They concluded that this skeleton was deposited in at least 100 m water
depth, but regardless – this specimen was certainly deposited on the
Pleistocene shelf (…because the specimen was found 72 m above sea level).
Modern studies of whale falls have demonstrated that Osedax has a
difficult time colonizing carcasses on the continental shelf, owing to
intermittent currents and sedimentation events. However, Anderung et al.
speculate that rapid sedimentation caused by retreating ice pack along the
saline front (where clay flocculation is accelerated). This is difficult to test,
and to be fair – perhaps 1 out of 1,000 Neogene marine mammal fossils I have
ever seen have convincing evidence of Osedax bioerosion, probably
resultant from their deposition on the shelf – and I think this is a more
likely reason for the lack of Osedax colonization.


Fossil ambergris from the Pleistocene of Italy.
One of the most fascinating taphonomic discoveries in years
was the report of fossil ambergris from the early Pleistocene of Italy.
Numerous large coprolites – a few dozen bearing cephalopod beaks and hooklets –
were found in Pleistocene muddy strata. Admittedly, this was pretty unexpected
– ambergris is known to be positively buoyant (although, admittedly – the
buoyant examples are those we see – who knows what proportion sinks rather than
floating) – and there is currently debate regarding how hard it is when it is
inside the digestive tract; it is thought by some that while in the body it
remains somewhat ‘compliant’ and only hardens after floating at sea for
extended periods of time. Ambergris is only known to occur in giant sperm whale
guts (Physeter macrocephalus), although in this case no cetacean body
fossils exist to corroborate such an association. I've already discussed this exciting find.

Skeleton of the holotype of Neoparadoxia cecilialina.
Barnes, L.G. 2013. A new genus and species of Late Miocene
paleoparadoxiid (Mammalia, Desmostylia) from California.
Natural History
Museum of Los
Angeles County
Contributions in Science
521:51-114. Direct link to pdf.
One of the studies published this year that is more
geographically relevant to this blog is the new monograph on desmostylians by
Larry Barnes (LACM). The main fossil in the paper is a new, absolutely gorgeous
paleoparadoxiid from the Monterey Formation – this specimen includes a complete
skull, mandibles, complete vertebral column, and appendicular elements. Barnes
named this as a new genus – Neoparadoxia cecilialina. I had heard ahead
of the publication that it would be named as a new genus, and I was unsure of
the rationale prior to reading the paper. A quick skim of the paper revealed no
solid rationale for splitting the genus Paleoparadoxia into three,
cladistic or otherwise: all four species – the new species, and formerly P.
repenningi, P. weltoni, and P. tabatai – all form a
monophyletic group in Barnes’ cladistic analysis anyway, which Barnes named the
Paleoparadoxiinae. Why not just keep things simpler and leave it as one genus?
I haven’t read the entire monograph, so I’m not quite ready to judge whether or
not this is a case of oversplitting. In addition to erecting a new genus name,
Barnes also referred the Stanford skeleton – Paleoparadoxia repenningi –
to the new genus, recombining it as Neoparadoxia repenningi. He also
established a new genus for the latest Oligocene-earliest Miocene
paleopardoxiid from the Skooner Gulch Formation, recombining it as Archaeoparadoxia
weltoni. The paper includes many dozens of beautiful figures and
illustrations, and certainly appears to be one of Barnes’ more significant
contributions to our field. I think I’ll have to cover this study with a
dedicated blog post in the future, after I have more time to read and digest
the paper.


Phlogenetic and geographic placement of the Tunisian sea cow specimen.
Archaic sirenians are probably not my strong point. The only
sirenians I’ve ever dug up are all hydrodamalines – which represent, I would
say, the tragic pinnacle of sirenian evolution. So, as a result of reading
mostly about crown sirenians, I’ve not strayed much into the literature on
archaic sirenians – so this paper was a bit new to me. The problem with
sirenians is that they are members of the Afrotheria – a group of mammals with
African origins – but the earliest and most primitive known sirenian is Prorastomus
from the early-middle Eocene of Jamaica. So, we’re missing the earliest
representatives of the group from where they should be – Africa.
The earliest known and most primitive sirenians – Prorastomus, Pezosiren
– are quadrupedal sirenians that had not yet lost their hindlimbs (Pezosiren
in particular, in terms of its phylogenetic placement and terrestriality, is a
sirenian analog of Pakicetus). These two sirenians from Jamaica
have been found in estuarine sediments. Benoit et al. report a new sirenian
earbone from the late early Eocene (late Ypresian to early Bartonian) of Tunisia,
in a lacustrine limestone. The anatomy of this earbone is relatively more
primitive than Prorastomus from Jamaica,
making the Tunisian specimen the most primitive known sirenian. Unfortunately,
the new specimen is far too incomplete to be named – but the earbones of
mammals – particularly of cetaceans – are very informative and isolated
earbones such as this can often be accurately identified (for example, in some
morphological cladistic analyses, up to 1/3 of all phylogenetic characters are
periotic or tympanic bulla characters). In this study, Benoit et al. were able
to conduct a phylogenetic analysis, corroborating the stem position of this
taxon. They suggest that pachyostosis of the periotic is tied to aquatic
adaptation in this early sirenian (as in other sirenians and cetaceans).
Another study by Mark Clementz on isotopes suggested that sirenians took a
direct route to marine life, instead of having a prolonged intermediate
freshwater aquatic stage. The presence of the Tunisian specimen in freshwater
settings, however, suggests that sirenians may have indeed adapted to a transitional
freshwater environment before leaving Africa. On a
humorous parting note, this paper has to have the record for the number of
authors per single fossil bone in paleo literature.

Skull of the extremely strange ziphiid Globicetus from offshore Portugal.
In 2007, Giovanni Bianucci, Olivier Lambert, and Klaas Post
published a large number of fossil beaked whales (Ziphiidae) trawled from
offshore South Africa,
which included multiple strange new genera and species. Most of these were
found inside phosphorite nodules on the seafloor, and unfortunately because of
this no detailed age data is known.
A new study published this year Is sort of a followup to
that South African study, and reports many new – and also strange – ziphiids
from offshore Portugal.
These include a new species of Tusciziphius, known from Portugal and
also South Carolina – that has a large ossification formed as a
posteriorly-rising “fin” of bone on the rostrum, somewhat similar to Aporotus
(although in Aporotus it is formed by the paired premaxillae and is
transversely bilobed). Several new skulls are referred to Tusciziphius
atlanticus. Another strange ziphiid, Imocetus, bears a strangely
elongate facial region and a rostrum with a very wide, triangular base, vaguely
reminiscent of the overall shape of a beluga skull in dorsal view; however, it
bears elongate maxillary ridges, and a pair of short conical crests on the base
of the rostrum. Other ziphiids include new records of Caviziphius, Ziphirostrum,
and a new species of Choneziphius. Arguably the strangest new cetacean,
however, is Globicetus – a robust beaked whale with a bizarre
football-shaped ossification on the middle of the rostrum. I remember at the
2011 Aquatic Tetrapods conference in San Diego,
Ted Cranford asked if the spherical structure could be a fossilized melon –
unfortunately the chances of that happening are remarkably slim, although some
soft tissue structures such as baleen and intervertebral disks have been
preserved in Peruvian mysticetes. The structure is actually bone, and is formed
by the medially fused premaxillae. Many ziphiids bear strange rostral
ossifications and crests, and they probably have some function in reflecting
sound (air is an effective acoustic barrier, but bone makes for a close
second); the ossifications appear to be distinctive amongst various species, and
are also known to arise via different developmental means (a paper by Olivier
Lambert in Comptes Rendus Palevol reviews this nicely, and would be a good
topic to cover on here at some point). In a way, Globicetus isn’t really
any stranger than extant Hyperoodon (bottlenose whales), which has a
pair of tall maxillary crests on either side of the melon; it’s just that Hyperoodon
is “familiar” but strange, and Globicetus is “unfamiliar” and
strange, so to speak.

Fossil platanistine periotic from Amazonian Peru.
This short communication presents a pretty surprising
discovery, even if it’s just a single earbone: a record of a Platanista-like
dolphin (identified to Platanistinae) from the Peruvian Amazon. Most of us
probably think of the Pisco-Ica region on the Pacific side, which is dry and
dusty and effectively a big desert; however, it’s easy to forget about the part
of Peru on the
other side of the Andes, which is technically part of Amazonia.
A single periotic was collected from late middle Miocene fluvial rocks here,
and the periotic shares numerous features in common with extant Ganges
river dolphin, Platanista gangetica. For the geographically
uninformed, this may not seem like anything huge: the Ganges
river is in India,
and Platanista is currently relegated to Pakistan,
India, and Bangladesh,
and is totally riverine in distribution. Although G.G. Simpson placed all river
dolphins into a single subfamily Platanistoidea, it now appears that each river
dolphin really needs its own family: Pontoporiidae (Franciscana/La Plata river
dolphin), Iniidae (Amazon river dolphin/Boto), Lipotidae (Chinese river
dolphin/Baiji), and Platanistidae (Ganges river dolphin/Susu), as these occur
as a paraphyletic group in most molecular and many morphological cladistic analyses
of odontocete relationships. So, this new record indicates that three of these
families – Pontoporiidae, Iniidae, and Platanistidae – were all swimming around
in coastal waters or rivers of South America during the
Miocene. This has some curious biogeographic implications; it’s thought that
these reflect independent adaptation to riverine life amongst archaic
odontocetes, with many of their oceanic relatives going extinct and being
replaced (passively or competitively) by more recently evolved delphinoids.
This new find demonstrates that platanistines were not restricted to Asia,
and an early Miocene specimen from Oregon
indicates they were not uniformly riverine during the Miocene. Bianucci et al.
hypothesized that platanistines had a marine, North Pacific origin, followed by
invasion of rivers in South America and southern Asia,
and subsequently going extinct in South America.
Bianucci et al. also suggested that the long rostra of “platanistoids” (sensu
Simpson) was a preadaptation for freshwater existence – perhaps suggesting
a limited ability of delphinoids to colonize freshwater environments (although
plenty of freshwater brevirostrine delphinoids exist: Orcaella spp., and
Neophocaena, and a couple of other more longirostrine delphinids).


Holotype skull of the new Pliocene dolphin Septidelphis.
Giovanni Bianucci’s Ph.D. thesis and eventual publications
in the late 1990’s dealt with a comprehensive reassessment of the Pliocene
odontocete assemblage from Italy,
which is a pretty sizeable assemblage and certainly no small feat. Of many of
the historical specimens, there was one he was never able to track down and
examine: a specimen of Stenella cf. frontalis reported in 1980;
unfortunately the author of the study did not report where the specimen was
reposited, and died before Bianucci could contact him about it. After recently
learning the whereabouts of the specimen, Bianucci examined and reevaluated the
specimen. Owing to some cranial differences, he named it Septidelphis morii –
one peculiar feature is the presence of a fairly wide mesorostral groove, which
is quite a bit wider than in other delphinids (elegantly shown graphically
within the paper). Instead of conducting a dedicated morphological cladistic
analysis of small bodied delphinine delphinids – no doubt a difficult
undertaking thanks to the skulls of modern delphinines looking damned near
identical (if I tried it, it would surely result in tears and hair being pulled
out) – Bianucci used a molecular “scaffold”, constraining the tree topology and
phylogenetic position of extant delphinids based on their occurrence in
molecular phylogenies. 14 morphological characters were used to place the
fossil taxa on the tree, with Pliocene fossil delphinids Astadelphis, Etruridelphis,
and Septidelphis forming a paraphyletic stem group of delphinines. Each
of these taxa were formerly identified as fossil Stenella; Bianucci
suggests that, based on his results and the molecular phylogenetic results of
others, that extant genera of delphinids probably did not arise until the
Pleistocene, and that the majority of Pliocene delphinids are probably better
attributed to extinct genera. Interestingly, given the recent proposal by
William Perrin and others to combine Delphinus, Tursiops, Stenella,
Lagenodelphis, and Sousa under Delphinus – which would no
doubt make all of these extinct delphinine genera from Italy
Delphinus as well. Is this a case – modern and fossil species alike - of
taxonomic oversplitting, with recombining everything as Delphinus as the
solution? I’m not sure, and certainly more morphological work on delphinines is
necessary. To be fair, I have a hard time swallowing the idea that extant
delphinid genera were not present at all during the Pliocene, but that’s just a
gut opinion.

During the late 19th century, a number of fossil
baleen whales were named from the Miocene and Pliocene of Belgium. Many of
these are of debatable utility and diagnosability, and many have been
demonstrated to be chimaeras – collections of isolated, non-associated bones
which the author interpreted to belong together. One of these fragmentary
baleen whales was described as Isocetus depauwi – it includes a series
of cervical and thoracic vertebrae, a partial mandible, a tympanic bulla, and
an ulna. Reevaluation of the type specimen indicated that it is indeed a
distinct genus and species, albeit difficult to compare with more complete
fossil mysticetes; a nearly complete skull, periotic, tympanic bulla, mandible,
and vertebrae was referred by the German paleontologist Abel to Isocetus
depauwi. This specimen was described in detail and figured by Bisconti et
al., who named it as a new species of Parietobalaena – Parietobalaena
campiniana. Interestingly, this study proposed some of the first
synapomorphies for the genus Parietobalaena, including numerous earbone
features. Two poorly known mysticetes which were transferred to Parietobalaena
by Steeman in 2010 – Heterocetus affinus and Idiocetus laxatus –
were concluded to likely be valid genera owing to periotic differences. Most
significantly, a new clade of mysticetes was named: it includes all mysticetes
crownward of right whales (Balaenidae), including Balaenopteridae (rorquals),
Eschrichtiidae (gray whales), Cetotheriidae sensu stricto (true
cetotheres), and cetotheres sensu lato, jokingly referred to as
kelloggitheres (e.g. Kellogg’s cetotheres – Parietobalaena, Diorocetus,
Aglaocetus, Thinocetus, Halicetus, Pelocetus,
etc.). Admittedly, kelloggitheres are my least favorite group of mysticetes –
they all look the same to me. There’s only one problem with the proposal of
this new taxon – depending upon which phylogenetic result is achieved, it may
or may not be equivalent (or mutually exclusive) with Balaenopteroidea, sensu
Deméré et al. (2005). In Deméré et al. (2005), Cetotheriidae ss. appear on the
stem relative to balaenids, but in this study, cetotheriids occur crownward of
balaenids; in Deméré et al., everything crownward of balaenids belongs to
Balaenopteroidea, whereas everything crownward of Balaenidae in this new study
is termed Thalassotherii. By the way, I was pretty pleased and entertained to
hear the term kelloggithere used in at least one SVP talk this year.


The Pliocene marine vertebrate assemblage from the San Gregorio section
of the Purisima Formation.
Boessenecker, R. 2013. A new marine vertebrate assemblage
from the Late Neogene Purisima Formation in central California,
part II: Pinnipeds and Cetaceans. Geodiversitas 35:815-940.
This new study which came out on Dec. 27 was almost in time
for Christmas. It marks the culmination of 8 of research on my part. In 2004 I
received a tip to check out some bones at a beach by a local surfer, and when I
got out there I found numerous bonebeds and hundreds of bones sticking out of
the cliffs. It was my first summer home from college in Montana,
and I had an idea for fieldwork – virtually nothing had been published on
fossil marine mammals from the Purisima Formation, and literally nothing had
been published on material outside the Santa Cruz
area – so this was a ripe piece of fruit to pick. I received my first permit in
2005, had it renewed for 2006, and received another permit for 2010 to 2011.
During these periods I collected a rather large number of marine vertebrate
fossils, which would eventually represent over thirty species of sharks, bony
fish, marine birds, pinnipeds, and cetaceans. Two earlier papers published all
of the sharks, bony fish, and birds, including the embarrassingly large
pelagornithid humerus which Adam Smith and I published in 2011 (the drawer
label at UCMP is titled “Bobby’s big bird”). This new paper is concerned with
the fossil marine mammals, and includes three pinnipeds, several baleen whales
including Eubalaena (right whale), Herpetocetus, and a new
species of Balaenoptera – Balaenoptera bertae, which I’ve named
in honor of Dr. Annalisa Berta. Odontocetes include delphinids, a
globicephaline, an indeterminate sperm whale, Parapontoporia sternbergi,
and several phocoenids including the strange “skimmer porpoise” as well as a
harbor porpoise like fossil I’ve identified as cf. Phocoena. In addition
to reporting all of these marine mammals, the paper includes a review of
various Pliocene marine mammal assemblages from around the globe.
Interestingly, I found that when I tallied up the proportion of extant to
extinct genera in various assemblages, Pliocene marine mammal assemblages from
the Pacific consistently had fewer extant genera than the North
Atlantic. The Yorktown Formation of the eastern US, for example,
is approximately 55% extant, whereas it is about 30-35% or so for the eastern
North Pacific, and about 17% for the eastern South Pacific. This suggests that
many extant marine mammals appeared earlier in the Atlantic
than they did in the Pacific. Furthermore, many Pliocene assemblages seem to
have all sorts of non-cosmopolitan “bizarre” species without modern ecological
analogs in the region. All of these data, in concert with a Pleistocene marine
mammal assemblage that is mostly comprised of extant taxa, indicate that a
higher degree of faunal provinciality prevailed during the Pliocene and that
various extinctions occurred amongst these groups sometime during the early
Pleistocene.

Revised stratigraphic range of Herpetocetus with some other charismatic megafauna added for context.
This study published in Naturwissenschaften early this year reports a surprisingly late surviving example of the archaic mysticete Herpetocetus - the specimen may be as young as 700 thousand years. This has some interesting implications for how we think about marine mammal faunal change during the Pleistocene. Read about it here.

A collage initially intended for a cover image - barnacle encrusted sea lion bones and their locality.
This paper published earlier in the year in Journal of Paleontology is another taphonomic contribution of mine, and reports a new occurrence of barnacle encrusted marine mammal bones - and expands on the possible data we can squeeze out of the fossil record from encrusting invertebrates. Read about it here.


Life restoration of the extinct "killer" walrus Pelagiarctos.
This paper by myself and Morgan Churchill describes a new fossil of Pelagiarctos and reevaluates the problematic "killer walrus" hypothesis for the original Pelagiarctos thomasi material from Sharktooth Hill. Read more about it here, here, here, here, and here.

Life restoration of a globicephaline whale from the Purisima Formation.
This paper describes new records of globicephaline whales from the Purisima Formation of central/northern California. Read more about it here.

The holotype skull of the new beaked whale Notoziphius from Argentina.
This is one of the few papers published this year which I
had the fortune to review. This study reports a new genus and species of beaked
whale from the late Miocene of Argentina, collected by (late) local fossil
collector Rodolfo Brunet. The specimen includes a relatively large skull,
approximately Mesoplodon sized – missing the anterior part of the
rostrum, and also some mandibular fragments. Monica Buono and Mario Cozzuol
named this new genus Notoziphius, and named it after the collector – Notoziphius
bruneti. Phylogenetic analysis indicates that this new ziphiid forms a
clade with Messapicetus, Ziphirostrum, Beneziphius, and Aporotus.
This new beaked whale is the first record of a fossil ziphiid from the
southwestern Atlantic, and demonstrates that ziphiids
were widely dispersed and diverse during the Miocene.


Fossil pygmy right whale mandible from the late Miocene of Argentina.
This new paper continues last year’s Caperea frenzy:
this is the third fossil neobalaenid/neobalaenine to be reported in the last 16
months: the paper reports a partial mandible from the late Miocene of Argentina
that is most similar to that of the extant pygmy right whale, Caperea
marginata. It notably differs in having a larger coronoid process (reduced
to absent in adult, modern Caperea, but retained in juveniles – see Marx
et al., below), and in having a slightly less dorsally arched mandible; however,
the features preserved are unique to neobalaenines. This specimen is the oldest
known pygmy right whale, at approximately 9-10 million years old; it is about
3-4 million years older than the recently described Miocaperea from Peru.
These attest to an old origin for pygmy right whales, and this specimen is just
slightly younger, for example, than the oldest well-demonstrated fossil
balaenopterids. Fossil balaenids, on the other hand, extend back to the early
Miocene (~10 Ma).


Fossil skull of an undescribed kekenodontid from the Oligocene of New Zealand, currently being studied by labmate Josh Corrie.
Although published in 2013, this paper came out “online
early” in 2012 and I covered it in last year’s post, which you can read here.

The strange taxonomic and phylogenetic framework for sharks in Diedrich 2013.
This paper by Cajus Diedrich is a followup to the 2011 paper
on a supposed pinniped from the Eocene (a highly unlikely and poorly supported
case which I summarized on this blog a couple years ago). This new paper
reports the co-occurrence of the aforementioned “pinniped” in addition to a
purported Protosiren rib, and a couple of bone fragments questionably
identified as protocetid remains (which had been more appropriately identified
by Diedrich as indeterminate mammalian remains in a previous paper); rationale
for these identifications is not provided in the paper (which admittedly is not
a solid foundation). Curiously, he uses a rather bizarre taxonomic framework
for sharks, concluding – again without evidence – that the Carcharodon
carcharias lineage can be traced back to the Eocene, and that both Carcharodon
and Carcharocles were separate entities during the Eocene. The teeth
identified as Carcharodon are clearly just small Carcharocles teeth
that have been misidentified; the earliest taxon that can be reliably ascribed
to Carcharodon is Carcharodon hastalis in the Miocene, as shown
by Dana Ehret’s careful work. Diedrich argues that the co-occurrence of three
marine mammals, and the earliest appearance of serrated Carcharodon and Carcharocles
– is indirect evidence that the latter evolved after marine mammals first
appeared in the North Sea. Is correlation = causation in
this case, assuming for one moment the marine mammals are correctly identified
(or even identifiable)? Or does the co-occurrence of these specimens in a time
averaged, taphonomically concentrated horizon instead indicate a physical
control on their occurrence simply by virtue of these sharing a calcium
phosphate mineralogy which may be concentrated in certain sedimentological
circumstances such as this? From a taphonomic perspective, any claim that there
is a causal link is a bit of a stretch. It’s not really clear what the science
was here aside from storytelling, and the puzzling taxonomic framework for
sharks and lack of defense for marine mammal identifications undercuts this
paper’s significance.
Full disclosure: I reviewed this paper twice for two
different journals, and was more than a little surprised to see it published –
effectively unabridged replete with all of its original typos – in a third
journal, less than a month after it was rejected from the second journal.
Despite not utilizing a shred of my constructive review comments, I was the
only reviewer thanked in the published paper…


Hand-drawn speculative cladogram of diphyletic sirenian evolution by Diedrich.
This new study by Cajus Diedrich is a followup to the above
study and the 2011 paper on a very dubious report of an Eocene pinniped fossil,
and purports to document a new record of Protosiren from the proto-North
Sea, and demonstrate evidence that sirenians are biphyletic (not monophyletic,
and had two separate origins and is therefore not a biologically real group).
The new fossil of Protosiren is just a rib – although ribs are
diagnostic for sirenians as a whole, they are not exactly a rich treasure trove
of morphological features, and I’m skeptical that such an identification is
based on well-founded reasoning. Apparently Protosiren does have
distinctive bone histology (thanks to Jorge Velez-Juarbe for the heads up), but
no attempt at sectioning the specimen was made (in fact, the fossil remains in
a private collection – which is not so good). Little morphological evidence is
marshaled to support the identification as Protosiren. The central tenet
of the paper – the identity of the main fossil reported within – is not solidly
founded, in other words. The rationale behind the other point of the article –
is not clearly accessible in the article – no attempt at conducting a
phylogenetic analysis which reflects sirenian biphyly is made, although a
pretty diagram showing sirenian diphyly is included. Importantly, no cladistic
analysis was executed or reported upon – it is necessary to note here that the
reported tree is a hand-drawn cladogram. That’s fine, it’s still a hypothesis,
but not one that was tested by the author. It’s unclear after reading just
where the “science” was in this study.


Cluster diagram of various fossil and modern marine and terrestrial mammals, grouped together based on microwear data.
This new paper by Julia Fahlke and colleagues investigates
dental microwear on the teeth of archaeocete whales to document changes in diet
during the land to sea transition in early cetaceans. Fahlke et al.
investigated a pakicetid, several protocetids, and basilosaurids – fully
bridging the terrestrial to marine transition. The habitat and locomotion of
archaeocetes are already well known thanks to functional and isotopic studies,
but it’s unclear exactly what archaeocetes were eating. For example: did
aquatic feeding or adaptation for aquatic locomotion occur earlier in whale
evolution? Or vice versa? The study of microwear is the microscopic analysis of
damage to enamel, which is broadly correlated with diet. In extant herbivorous
mammals, the number of pits and scratches is tied to how much silica is
ingested and chewed – often in the form of phytoliths that grow in grass, and
information like this can tell you whether or not a mammal was a browser or
grazer. Dental microwear has historically been applied more frequently to
herbivorous mammals, but recently a number of studies by Brian Beatty and others have applied
microwear to marine reptiles in order to evaluate feeding ecology – so there is
scope for applying these methods to toothed marine mammals (sorry,
chaeomysticetes). As it turns out, pakicetids and semiaquatic (pinniped
equivalent semiaquatic-ness) protocetids share similar microwear patterns,
suggesting that pakicetids were indeed feeding in the water. Basilosaurids, on
the other hand, were found to have microwear indicating feeding on occasional
harder prey – marine mammals, sea birds, etc., in addition to exhibiting
extensive tooth damage related to contact with the bones of prey items. One
protocetid, Qaisracetus, had microwear patterns similar to hyenas and
killer whales, suggesting a large component of warm-blooded prey in its diet
(e.g. other marine mammals, sea birds, large fish, etc.). In summary, they
concluded that specialized piscivory and teuthophagy in extant odontocetes is
derived from what was formerly a much more generalized diet during the Eocene –
perhaps permitted by a dentition that could still shear, as opposed to modern
odontocetes which predominantly swallow prey whole (killer whales being an
exception).


Fossil sea cow as discovered in 1975 in a cave in New Guinea.
It’s always difficult finding vertebrate fossils in the
tropics; carbonate rocks predominate in equatorial latitudes, and are not a
great environment for preserving abundant marine vertebrate skeletal material –
coastal carbonate systems tend to be characterized by relatively rapid,
aggradational deposition, as opposed to vertebrate-rich low subsidence settings
in siliciclastic basins where bones can be concentrated quite densely. As such,
the marine vertebrate fossil record from Oceania –
outside Australia
and New Zealand
– is quite limited. Occasionally, interesting gems are found though. A caving
expedition in the mid 1970’s came across a series of bones in early-middle
Miocene limestone in New Guinea
(admittedly, not a very small island). As it turns out, this partial skeleton
belongs to a small but indeterminate sirenian (sea cow). The extant Dugong currently
inhabits Australasian, Indonesian, and Philippine waters, but has a crappy
fossil record- it’s known from various Holocene localities in Australia
and New Guinea,
but only scraps of sirenians are known from earlier rocks in Australia.
Eocene sirenians are plentiful further north and west in the Indian
subcontinent as well as Java, but little evidence can be marshaled even to say
that sirenians were present (or absent) during the intervening time. This new
discovery suggests that sirenians indeed probably inhabited this region, and
fossils of them from rocks of this time have so far gone undiscovered. Fitzgerald
et al. suggest that this is due to incomplete sampling, and that other rocks in
Australia may
yet yield additional sirenian material.


This fascinating new study utilized ancient DNA from relatively young
fossils of North Atlantic balaenids – bowhead whales (Balaena mysticetus) and right whales
(Eubalaena glacialis) to examine evolutionary dynamics of the two whales
during the late Pleistocene. They used about 44 samples from subarctic and cold
temperate latitudes in the North Sea – United Kingdom, Denmark, Sweden, and Norway. Specimens of late Pleistocene age turned out to all represent Balaena
mysticetus, while those from the same latitudes – but from the Holocene –
all represented Eubalaena glacialis. Currently, Balaena mysticetus is an
arctic mysticete and is tied to ice-bound regions, whereas Eubalaena –
all three putative species – are temperate water whales. The combination of
identity (based on ancient DNA) and radiocarbon dates, in concert with paleoclimate suggests that
during the last glacial period of the late Pleistocene, bowhead whales
inhabited latitudes much further south than at present. During the Holocene, as
icepack receded North and the earth recovered from the last glacial maximum,
warmer water (note: still cold temperate and temperate – warmer, but
still not “warm” or warm temperate) Eubalaena replaced the bowhead whale
in those latitudes. This is a rather nice demonstration of habitat tracking in
the fossil record of an extant mysticete.

This short article reports a
partial odontocete rostrum with an associated tooth that shows evidence of
tooth disease. The tooth has a large bulge in the root below the base of the
crown. The bulge is not actually part of the tooth, and the morphology,
microstructure, and chemistry of the substance is consistent with it being a
deposit of dental calculus – in vernacular language, plaque. This condition
indicates that the dolphin likely had periodontal disease. Although periodontal
disease has been reported in several extant odontocetes, this is the first known
case in a fossil odontocete.

Location and radiocarbon ages of fossil and subfossil Eubalaena (purple) and Balaena (blue) occurrences in the eastern North Atlantic.

The novel phylogenetic hypothesis of Caperea relationships.
Fordyce, E., and F. Marx. 2013. The pygmy right whale Caperea
marginata: the last of the cetotheres. Proceedings of the Royal Society B 280: 20122645.
Another paper from Caperea fever. This new paper by
my adviser R. Ewan Fordyce and former labmate Felix Marx presents a provocative
new hypothesis on the phylogenetic relationships of the pygmy right whale,
positing that it is most closely related to extinct herpetocetine cetotheriids
such as Herpetocetus and Nannocetus, which resurrects the
Cetotheriidae from extinction. I’ve already covered this paper here.

Morphological changes in cetacea. There's a lot of labels here, so you can go ahead and consult the actual paper.
This large study is a review paper of sorts, and reviews
various aspects of molecular and morphologic evolution of cetaceans. The study
does reanalyze molecular and morphological data, but this is one of the more
robust and up to date reviews of cetacean evolution and nicely summarizes
current ideas on changes in locomotion, feeding ecology, sensory biology, sound
production, brain size, diving adaptations, respiration, habitat (e.g.
terrestrial v. aquatic v. marine), and other various aspects. There’s really
too much to cover here, but if you’re looking for an excellent review of
cetacean evolution within a phylogenetic framework, peppered with beautiful
artwork by Carl Buell thrown to boot – look no further than this paper.


Partial skull attributed to Stromerius by Gingerich.
Historically collected specimens can often be problematic,
even if they have been cited and discussed by various earlier researchers.
There can be many changes in schools of thought and practice within a
discipline over a century or so; as a result, many early specimens – types or
otherwise – have a dramatically changed meaning to us relative to Victorian
scientists. A new paper by Phil Gingerich illustrates quite nicely why old
museum labels should never be trusted at face value. A skull and mandible of an
archaeocete at the Field Museum
in Chicago was historically
identified as Prozeuglodon osiris (=Saghacetus osiris of current
usage). It was sold to the Field Museum
in 1914 or 1915 by Richard Markgraf. However, detailed study by Gingerich
indicated that the two specimens differ in preservation and appear to represent
two differently sized individuals. The skull is too small to represent Saghacetus
or Dorudon, and Gingerich tentatively referred the specimen to the
smaller basilosaurid Stromerius (the type specimen of which is
represented by a vertebral column – albeit smaller than other basilosaurids).
The mandible is in fact referable to Dorudon atrox. Gingerich points out
that Richard Markgraf was not a paleontologist or a stratigrapher, and
highlights problems with accepting museum labels at face value. This issue is
familiar to me, and was hammered into my brain during the first few years of
research, and as a result of questioning previous identifications while
visiting various west coast museum collections, I developed a photographic and
mental atlas of Neogene marine mammal fossils from UCMP, SDNHM, LACM, and
others to construct a body of knowledge that I could use to evaluate and reidentify
museum specimens (and, my own collected material).


Reconstructed olfactory apparatus of extinct protocetid archaeocete.
One of the most fascinating papers in marine mammal
functional morphology this year was a paper by Stephen Godfrey, Erich
Fitzgerald, and Jonathan Geisler on olfaction in archaeocetes and mysticetes.
This study examined a fragmentary archaeocete skull from Virginia,
mostly consisting of a frontal shield; they reasonably make the case that the
specimen represents a protocetid rather than a basilosaurid. The specimen was
found by a private collector who has been really generous in donating material
from his collection to various institutions; it was found on a riverbank, with
no adhering matrix, although its protocetid morphology suggests it is Eocene
and may have been derived from the Piney Point Formation. The specimen includes
well preserved olfactory structures. The olfactory apparatus of this protocetid
is well-developed, in contrast to extant odontocetes – which appear to lack
many of the structures found in this archaic whale and other “macrosmatic”
mammals (macrosmatic means mammals with well-developed olfactory senses, such
as dogs); odontocetes are widely considered to lack a sense of smell. The
protocetid appears to have a well-developed sense of smell, probably far more
sensitive than in humans. Interestingly, sectioned skulls of modern minke
whales have an olfactory apparatus that is strikingly similar and functionally
identical to the Eocene protocetid, leading Godfrey et al. to state that the
protocetid had effectively modern olfactory anatomy. While it is clear that
well-developed olfaction is probably primitively retained in some modern
mysticetes (possibly all) and lost in odontocetes, the discovery of probable
well-developed olfaction in mysticetes is somewhat surprising, given that when
diving and foraging, the nostrils of cetaceans are closed off by the nasal
plugs so as to prevent ingestion of water (and drowning). Godfrey et al.
indicate that krill give off a particular odor, and it has recently been
proposed that bowhead whales use their retained sense of smell to identify
other whales, and locate “clouds” of planktonic prey; they suggest that
olfaction – when the whales are at the surface, of course – could be used for
finding dense accumulations of prey items.


Skull outlines of fossil squalodontids. The grayed area on the right hand side is the preserved portion of the partial skull described by Godfrey.
As a followup to the above article, Stephen Godfrey
published another great paper on olfaction – but this time in an archaic
odontocete – Squalodon, from the middle Miocene of Maryland. The
cross-sectional area of the olfactory epithelium is about 7 times than the area
of the ethmoid bone, whereas in most terrestrial mammals it’s about 16 times
larger; this demonstrates that (unsurprisingly) Squalodon had an
intermediate olfactory sense between archaeocetes and extant odontocetes – and,
that olfaction was probably gradually lost within odontocetes during the
Miocene.


Part of the holotype of the new basilosaurid Basilotritus uheni.
This new paper by Pavel Gol’Din and Evgenij Zvonok describes
a new genus and species of basilosaurid archaeocete from the late Middle Eocene
of Ukraine. This new fossil consists only of a partial tympanic bulla and
several vertebrae. The vertebrae are grossly pachyosteosclerotic and inflated
(vaguely resembling sirenian vertebrae), and bear a strange punctate texture on
the external bone surface. This skeleton was given the name Basilotritus
uheni, and named after archaeocete researcher Mark Uhen, and represents the
oldest fossil cetacean from Eastern Europe. Phylogenetic
analysis recovered Basilotritus uheni as an early diverging
basilosaurid. Interestingly, Gol’Din and Zvonok noted the similarity between
the vertebrae of Basilotritus uheni and “Eocetus” wardii
from the eastern USA,
and recombined the latter as Basilotritus wardii. Several other
fragmentary and problematic fossil cetaceans, such as Platyosphys, may
have something to do with this new genus.

Sperm whale teeth from the late Miocene of Moldova.
Gol’Din, P. and V. Marareskul. 2013. Miocene toothed whales
(Cetacea, Odontoceti) from the Dniester
Valley: the first record of sperm
whales (Physeteroidea) from the Eastern Europe. Vestnik
Zoologii 47:21-26.
This short study reports several isolated teeth from an
indeterminate physeteroid from the Tortonian (~9-12 Ma) of Moldova
(a small country sandwiched between Ukraine
and Romania).
Two teeth apparently representing the same taxon are similar to teeth of the
wastebasket taxon “Scaldicetus” in having a large enamel cap and swollen
roots, and another tooth with an elongate and narrow root looks a bit like some
“physeterines” (=Physeteridae of some workers), potentially indicating that two
sperm whales were present. In the grand scheme of things, these are admittedly
not very old, since there are plenty of examples of middle Miocene sperm whales
(e.g. Aulophyseter from California),
and Ferecetotherium from the Caucasus is
potentially late Oligocene. These are, however, the first records of
physeteroids from eastern Europe, a region which is becoming better known in
terms of cetacean fossils thanks to recent efforts by the author, Pavel
Gol’Din.


Skull and subantarctic locality of Africanacetus from the seafloor.
A second paper forms a followup to the 2007 paper on fossil
ziphiids trawled from th seafloor off South
Africa (This paper was covered last year as
it was evidently online early in 2012 and only published this year, but I’ve
typed up a longer and more fitting summary this year). This study by Pavel
Gol’Din and Karina Vishnyakova reports two partial skulls of the beaked whale Africanacetus
from offshore Antarctica, at a remarkably high latitude – approximately 60º
south, and about midway (longitudinally speaking) between Australia and South
Africa. The two skulls are slightly larger than the skulls reported from
offshore South Africa,
and differ in having a more well-developed mesorostral ossification of the
vomer, leading the authors to simply identify the skulls as Africanacetus
sp. These authors further hypothesize a circum-Antarctic distribution of Africanacetus,
and also note that it is so far the highest latitude fossil ziphiid yet known;
an interesting parallel is the Pliocene ziphiid-convergent delphinid Australodelphis
from Antarctica named by Ewan Fordyce about a decade
ago.

Articulated skeleton of the holotype of Cetotherium riabinini from the late Miocene of Ukraine. Scale bar = 1 meter.

Articulated skeleton of the holotype of Cetotherium riabinini from the late Miocene of Ukraine. Scale bar = 1 meter.
It’s been a good year for Pavel Gol’Din – this study, also
published this year in Acta Palaeontologica – reevaluates the skeletal anatomy
of Cetotherium riabinini, a well preserved cetotheriid sensu stricto
from the late Miocene of Paratethys (collected in Ukraine).
I had the pleasure of reviewing this article last fall – and actually conducted
my peer review during Hurricane Sandy when I was shuttered in my friend’s
brownstone apartment in Washington D.C. (I was unable to go into the
Smithsonian for about four days). Cetothrium riabinini is a more obscure
species of Cetotherium from Paratethys; the genus is based on Cetotherium
rathkei, which is known from a skull – but when Brandt described it
initially, there was apparently a very narrow and tapering maxilla, which I and
others did not really think was complete. As it turns out, the more complete
skull of Cetotherium riabinini indicates that the narrow rostrum was in
fact accurate as illustrated by Brandt. Cetotherium riabinini is small –
about four meters body length, with a tiny skull with an elongate rostrum. The
postcranial skeleton is well preserved, and includes Caperea-like platelike
pachyosteosclerotic ribs. The rostrum is “bent” slightly anteroventrally, and
it shares some peculiar features of the mandibular articulation with Herpetocetus.
Owing to some of these features, Gol’Din and others argued that Cetotherium
riabinini was adapted for benthic suction feeding much like today’s gray
whale (Eschrichtius robustus). On a similar note, an in press article by
Joe El Adli, Tom Deméré, and myself makes the same case for Herpetocetus
from the Pliocene of California.


Bony tumor in the late Miocene balaenopterid "Megaptera" hubachi.
This new paper by German colleague Oliver Hampe and others
is literally hot off the press, and came out just a couple of days ago in
Alcheringa. This paper is concerned with large bony protrusions on the
occipital shield of the skull of “Megaptera” hubachi, a fossil
baleen whale from the late Miocene of Chile which was initially thought to be a
fossil humpback whale relative. “Megaptera” miocaena is generally
plesiomorphic and shares various primitive characters with extant Megaptera,
explaining why the original author placed it in the same genus. Regardless, it
needs a new genus, as has been concluded by several researchers. The skull of “Megaptera”
miocaena has a strange bony lump on its occipital bone, in about the
same position as small tubercles in rorquals (Balaenoptera) and even
larger tubercles in the gray whale (Eschrichtius). In “Megaptera”
miocaena, it is only developed on the left side, whereas in gray whales
it is bilaterally symmetrical – which Hampe et al. indicate is likely evidence
that it is pathologic, and not a gray whale-like muscle attachment as suggested
by Michelangelo Bisconti. Hampe et al. identify the strange structure as a
benign bony tumor or osteoma – which apparently is the first known example of
this in cetaceans. The inside of the structure is homogeneous and very dense.
Previously reported pathologies identified as osteomas in extant cetaceans
reported that the structures were very porous, and Hampe et al. suggest that
these extant examples are probably not osteomas and represent some other type
of abnormal bone growth such as spondylitis.


Osteohistologic sections of fossil desmostylians.
Bone histology has been an excellent tool to gauge
aquatic-ness of fossil marine tetrapods; multiple groups have acquired extremely
dense bones upon invasion of the aquatic realm, thought to aid as ballast or to
modify trim (orientation while swimming). This study by my Japanese friend and
colleague Shoji Hayashi took postcranial bones of various desmostylian
specimens from Japan.
They found that the early desmostylian Ashoroa laticosta had
pachyosteosclerotic bones – that is, bones with a reduced medullary cavity and
an outwardly expanded cortex (e.g. inflated bones). The earlier diverging
desmostylian Behemotops, and Paleoparadoxa both showed evidence
of pachyostosis (inflated cortical bone). The most derived desmostylian, Desmostylus,
on the other hand, showed evidence of osteoporosis – decreased bone density
(the opposite of osteosclerosis). An increase in bone mass is tied to
hydrostatic buoyancy and body trim and correlated with inefficient swimmers.
However, decreased bone mass is related to hydrodynamic buoyancy control in
active swimmers. This trend parallels that seen in cetaceans and pinnipeds,
indicating all desmostylians have osteologic adaptations for aquatic life. Desmostylus
in particular appears to have been a more active swimmer and more adapted for
marine life than others – which does appear at odds with its inferred ecology
as a seagrass or kelp grazer (i.e. since aquatic plants and algae grow in the
photic zone and along the shoreline in shallow water). The spongy bone in Desmostylus
parallels most modern cetaceans, in addition to elephant seals.


Referred skull of Haborophocoena toyoshimai.
This paper reports a new specimen of the fossil porpoise Haborophocoena
toyoshimai. Haborophocoena is a porpoise that differs from extant
porpoises in retaining an asymmetrical skull: the right premaxilla is wider
than the left and extends further posterior to the left, in addition to the
right maxilla being wider than the left, and the vertex being offset to the
left side of the midline. A second species, Haborophocoena minutis, was
reported by the same authors in 2009 from a different locality – and from this
locality originated the new, second specimen of Haborophocoena toyoshimai.
The new specimen yields additional insights into the skull anatomy of this
porpoise.


Much of the fossil record of fossil
phocids consists of isolated bones and teeth. Partial skeletons are rare, but
known from the Pliocene of North America and Italy, and Miocene and Pliocene of
South America; fossils from other regions – northern Europe, Africa, Australia,
New Zealand, and the “Paratethyan” region (areas with deposits of the
Paratethyan sea, which effectively includes everything between southeastern
Europe (Romania, Hungary, Austria and east towards the Caucasus Peninsula –
e.g. Georgia, Dagestan). A series of publications by Irina Koretsky (Howard
University) have detailed fossil
phocid records from eastern Europe (primarily from Ukraine),
the Netherlands,
and the east coast of the USA.
A new study by Koretsky and student Sulman Rahmat describes and reevaluates
some robust phocid bones from the Middle and early Late Miocene of Ukraine.
They name two new species within the new genus Pachyphoca: P. ukrainica
and P. chapskii. They place this new genus within the subfamily
Cystophorinae, which purportedly includes modern hooded seals (Cystophora)
and elephant seals (Mirounga). However, this clade is a bit
controversial as work by Koretsky continuously refers to it in her work –
although it has not been recovered in a single molecular analysis. Instead, all
molecular analyses to date identify Cystophora as a phocine (closely
related to harbor seals, bearded seals, ribbon, and harp seals), and Mirounga
as a monachine (closely related to monk seals and Antarctic seals – e.g.
leopard, ross, crabeater, and weddell). Regardless, fossils reported in this
study definitely record the presence of a seal with peculiar
pachyosteosclerosis (thickened bones). Unfortunately, it does not appear that
these authors figured the holotype humerus of Pachyphoca ukrainica, as
that specimen does not appear in any of the figure captions. Interestingly,
this paper included “Afrophoca libyca” in its cladogram, a species that
would not be named until a separate paper was published later in JVP (early
2014), sort of making it a temporary nomen nudum.

Beautiful life restoration of the fossil ziphiid Ninoziphius from Peru.
Ninoziphius is an archaic beaked whale described by
Christian de Muizon in the early 1980’s from the early Pliocene Pisco Formation
of Peru. It was initially described in a brief article in French, and
subsequently a longer description was published as part of a monograph on Pisco
Fm. odontocetes – but again, in French. This new study redescribes the type
specimen in even more detail (and in English!), and reports new skulls which
preserve the vertex, which is damaged in the holotype. The cladistic analysis
in this study confirms that Ninoziphius is the most archaic known fossil
ziphiid. The feeding apparatus of Ninoziphius is less specialized for suction
feeding than extant ziphiids, owing to the retention of a homodont dentition
and elongate rostrum. Extensive tooth wear in Ninoziphius is interpreted
to correspond to benthic feeding, or capture of prey near the sea floor. Based
on facial cranial anatomy, Ninoziphius evidently was as capable of
echolocation as extant ziphiids; it also exhibited relatively enlarged pterygoid
sinuses, which appear to correspond to deep diving in ziphiids and physeteroids.
Despite all this, the vertebral column of Ninoziphius is more flexible
than extant ziphiids, with a longer cervical series; these suggest a less
stiffened vertebral column that is less well adapted to deep diving than extant
beaked whales.


The holotype skull of Brachydelphis jahuyaensis from the late Miocene of Peru.
This new study by Olivier Lambert and Christian de Muizon
reports a new species of pontoporiid dolphin from the Pisco Formation of Peru.
The dolphin is a new species of Brachydelphis. Brachydelphis mazeasi is
a short-snouted relative of the extant La Plata River Dolphin Pontoporia
blainvillei, notable for having an extremely short snout; it was described
in the late 1980’s by Christian de Muizon. This new species is somewhat younger
than B. mazeasi, and named Brachydelphis jahuayensis. Curiously,
it has a somewhat longer rostrum than the older species – although it is still
a notably short rostrum for a pontoporiid. If these two species belong in a
single lineage, it implies that this lineage developed a short rostrum from a
longirostrine ancestor (the primitive condition for pontoporiids), and
subsequently evolved towards having a longer rostrum.


The ascending process of the maxilla and coronoid process of the mandible in various mysticetes.
Yet another paper from Caperea mania. This paper
looked at juvenile and adult specimens of fossil and modern mysticetes to
examine the ontogenetic polarity of a few characters that influence the
phylogenetic position of the pygmy right whale. One feature is the lack of a
coronoid process in adult Caperea, which it shares with right and gray
whales to the exclusion of other mysticetes; however, juvenile specimens have a
triangular coronoid, whereas juvenile balaenids – and juveniles of archaic
balaenids – still lack one, suggesting that Caperea evolved from an
ancestor with a triangular coronoid. The other character, lack of an ascending
process of the maxilla, is shown to actually be present in juvenile Caperea,
but absent in juvenile balaenids. Again, this suggests that Caperea evolved
from an ancestor with an ascending process, which appears to have never been
present in balaenids. Both features then are not really synapomorphic, and
cloud our ability to effectively use them in phylogenetic analyses. As a bonus,
there’s a figure of a mandible and skull of Herpetocetus bramblei provided
by yours truly.

Study of cetacean brain size has been a bit controversial over the past
decade – back in 2006, a troubling study was published by anatomy professor Dr.
Paul Manger suggesting that modern dolphins weren’t actually very intelligent,
and instead had big brains for generation of heat – this rather unfortunate and
poorly informed study was quickly shot down by virtually everyone studying
dolphin intelligence, cetacean brain evolution, and cognition (all as coauthors
on one, massive takedown paper which I really ought to find time to discuss on
here someday). This new paper is a much, much more sober view, and examines
trends in EQ (Encephalization Quotient) through cetacean evolution. It’s
difficult to administer IQ tests to animals, primarily because they don’t speak
English (although communication and with some delphinids has been established
in some experimental settings). EQ instead assumes that larger brain mass
correlates with higher intelligence – it doesn’t always hold true, but on the
whole it is a pretty good indicator or predictor of animal intelligence. The
added benefit is that it is entirely anatomical – EQ can be assessed for dead
animals, as it is a measure of brain mass (or volume) relative to body mass.
Significantly, EQ can also be assessed in fossils if a reasonable body size
indicator can be found, and if a complete braincase is preserved and
CT-scanned. This study found that both body size and brain size tended to
increase through geologic time, and also that EQ decreases were common. For
example, in baleen whales body mass increased at a much faster rate than brain
size, resulting in the gigantism of today’s mysticetes – all of which have
relatively low EQ. In early Odontoceti, the opposite happened: the earliest
odontocetes were far smaller than basilosaurid archaeocetes, but brain size did
not decrease as quickly, and as a result EQ increased. Lastly, this study
points out that until only a few million years ago, the majority of mammals with
the highest EQ were all dolphins, not primates.


Periotics of the new albireonid from Japan (top) and Albireo whistleri from Cedros Island, Baja California (bottom).
Murakami, M., and Y. Koda. 2013. The first Pliocene
albireonid (Cetacea, Delphinoidea) periotic from the western North Pacific and
paleobiogeographic significance of fossil delphinoid ear bones of Na-arai
Formation of Choshi, Chiba, central
Japan. Japan
Cetology 23:13-20.

Mizuki Murakami and colleagues report on another Pliocene
porpoise (Phocoenidae) from Japan.
This one, unfortunately, is too incomplete to be identified or named, but
includes teeth, a strange rostrum, and quite a bit of the vertebral column. The
phocoenid was relatively small, with an abnormally narrow rostrum for a
phocoenid; sectioning of its teeth indicate that it was about four years old
when it died. The young age and skeletal maturity of this specimen suggest that
skeletal maturity was achieved quite early on during its ontogeny. The
vertebral column of this specimen is morphologically intermediate between that
of more primitive porpoises like Numatophocoena, and extant phocoenids
like Phocoenoides. This finding suggests that postcranial evolution
amongst phocoenids has been mosaic rather than gradual and ‘directed’.


Fossil gray whale mandible collected from the seafloor off Georgia.
This paper is a
followup to a Palaeontologia Electronica article by Garrison et al. (2012), and
reports on several fossil gray whale specimens recovered from the sea floor off
of Georgia (U.S.A.). Two gray whale specimens are represented by mandibles with
radiocarbon dates of about 30 Ka, and appear to represent juveniles, possibly
under one year old. These fossils demonstrate that at about 30 Ka, Eschrichtius
robustus was calving along the
eastern coast of North America, in addition to being the oldest known specimens
of the now-extinct North Atlantic population of gray whales (in the western
North Atlantic, anyway).


Restricted distribution of sirenians during the Pliocene in Europe and North Africa (yellow spots).
This new study summarizes the known fossil record of
sirenians (sea cows) in Europe and North
Africa. This region is now totally devoid of sirenians, although
they inhabited the Mediterranean from the Oligocene
through to the Pliocene. The disappearance of sirenians is an interesting
phenomenon, and one facet of late Neogene faunal change in marine mammal assemblages.
Prista et al. concluded that sirenians became extinct in the eastern North
Atlantic first, due to oceanic cooling and fragmentation of
seagrass habitats. Seagrass habitats were inferred to persist in the Mediterranean,
and extinction of Mediterranean sirenia was concluded to be caused by glacially
induced cooling.


Mandibles and measurements - gigantic jaws of Balaenoptera.
This study examined the mandibles of balaenopterid whales
(rorquals) to determine how they scaled with body size. They also point out
that the mandible of the blue whale is the largest vertebrate skeletal element;
perhaps an unremarkable finding, since blue whales are widely known to be the
world’s largest vertebrate animal, fossil or modern (some dinosaur fan boys
have proposed several sauropod dinosaurs that may be larger, but have produced
insufficient evidence to dethrone Balaenoptera musculus). They report
that a specimen of Balaenoptera musculus, USNM 268731, was from a 28
meter long female, and measure 6.8 meters in length. Damn, that’s huge. The
more interesting aspect of this study was their use of scaling relationships to
estimate body length from mandible size; many fossil balaenopterids are
incomplete, few with skeletons or any postcrania, although isolated mandibles
are abundant (they have a very high preservation potential owing to their large
size). Mandible length corresponds to skull length, and skull length correlates
well with body size in balaenopterids. They used three measurements – chord
length (i.e. straight line from tip to tip), curvilinear length, and condyle to
coronoid distance. They found that the relationship between mandibular length
and body length is nearly isometric, and also that condyle-coronoid length
decreases with increasing size. Although based on skeletal length and
mandibular measurements of extant mysticetes, they tested their mandibular
estimates with skull-based estimates for two fossil mysticetes with postcranial
skeletons. Mandibular estimates were comparable with skull-based estimates,
which is encouraging. Pyenson et al. then used two partial balaenopterid
mandibles from the Purisima Formation (my favorite rock unit) to give examples
of applying this method. The two mandibles from the Purisima Formation ended up
being reconstructed as 3.26 and 4.83 meters in length, total – smaller than
extant minke whales.


Physeteroid teeth from the Miocene of Spain.
This study reports some isolated teeth of a physeteroid
sperm whale from the Late Miocene of southern Spain.
They identify the teeth to the problematic genus Scaldicetus. Scaldicetus
is supposedly diagnosed by having robust teeth with primitively retained enamel
caps. However, teeth of the Scaldicetus morphotype have been found among
various skull morphologies – and teeth of this morphotype belong to several
extinct genera including Zygophyseter, Acrophyseter, Brygmophyseter,
and Livyatan. Scaldicetus as a taxonomic entity is virtually
meaningless. Admittedly, I only have a Spanish language version of the paper,
and am unable to read the rest of the work.


Map of fossil cetacean localities in Taiwan.
Tsai, C., Fordyce, R., Chang, C., and L. Lin. 2013. A review
and status of fossil cetacean research in Taiwan.
Taiwan Journal
of Biodiversity 15:113-124.
This new study by my labmate and fellow mysticete enthusiast
Cheng-Hsiu Tsai and colleagues summarizes current and former research on fossil
cetaceans from Taiwan.
This study is derived in part from Tsai’s master’s thesis – the other half of
which will be coming out in the Japanese journal Paleontological Research
sometime next year on fossil gray whales from the Penghu Channel (along the
western shore of Taiwan). Tsai et al. report the occurrence of numerous
mysticetes including balaenopterids, balaenids, and Eschrichtius (again,
to be considered in more depth in a following study) as well as delphinids, all
from Miocene through Pleistocene rocks. A fossil pilot whale (Globicephala
macrorhynchus) was trawled out of the Penghu Channel, and a well-preserved
globicephaline skeleton which was previously named Pseudorca yuanliensis
in an abstract-length publication. However, Tsai et al. pointed out that in the
absence of a description this name is a nomen nudum. They also indicate
that the taxon Balaenoptera taiwanica is based only on a tympanic bulla,
meaning that some caution needs to be exercised when using the name. Tsai et
al. conclude that although only a meager cetacean fossil record has been
established in Taiwan in contrast to Japan, there has been very little research
focus as well; preliminary field observations suggest a richer record than has
been published, and that much more field work needs to be done in order to
flesh out the cetacean fossil record of Taiwan.

This new paper by archaeocete
specialist Mark Uhen is an excellent, more or less comprehensive review with
commentary on the fossil record of Basilosauridae in North America (read:
United States, since I don’t think any archaeocetes are known from Canada or
Mexico, aside from a fragmentary cetacean vertebra from the Eocene of Vancouver
Island not positively identifiable as an archaeocete). The most well-known
basilosaurids from the US are of course Zygorhiza kochii, familiar to
most researchers studying Neoceti as it represents the de facto outgroup taxon
for comparisons and codings, and the king “reptile” itself, Basilosaurus
cetoides, the largest archaeocete. However, numerous less-well known
basilosaurids are summarized in the paper – the protocetid-like “Eocetus”
wardii, recombined as Basilotritus wardii in this new paper after
the above mentioned work by Gol’Din and Zvonok, Chrysocetus healeyorum,
a juvenile basilosaurid with large permanent teeth (suggesting monophyodonty as
in extant odontocetes) described by Mark Uhen and Phil Gingerich in 1998, the
ever problematic Dorudon serratus, also a juvenile, known by a
fragmentary skull with deciduous teeth, and Cynthiacetus maxwelli, named
by Mark in 2005 from a fragmentary large skull preserved with vertebrae not
anteroposteriorly elongated and shaped like soda cans - indicating a more “normal” cetacean than Basilosaurus
cetoides, although of similar skull size and morphology. This paper
highlights the taxonomic issues surrounding Zygorhiza – and recommends that the
well-preserved skull reported by Kellogg in his 1936 tome on the Archaeoceti (a
reference I use frequently) be designated as a neotype, since the Zygorhiza
kochii holotype is non-diagnostic. It bears stating that I have heard that
this is not necessary under the ICZN, and if I recall correctly a type specimen
need not be diagnostic; I also seem to remember that if a known holotype
exists, a neotype cannot be designated.

Pinniped localities in South America and fossil southern sea lion remains from Chile.
This new paper marks Anita Valenzuela-Toro’s publishing
debut, and describes several new pinniped fossils from the late Pliocene and
Pleistocene of Chile. Most are fragmentary, but nonetheless tell an important
story. The Pliocene specimens are a couple of ankle bones that are undoubtedly
those of extinct true seals, several genera of which (like Hadrokirus mentioned
above) inhabited the southeast Pacific during the Pliocene. However, the
Pleistocene specimens – probably late Pleistocene at that – are all otariid
specimens, most of which are identifiable to the extant South American sea
lion, Otaria byronia. The modern pinniped assemblage in South
America is entirely composed of otariids (Otaria, Arctocephalus)
in addition to the southern elephant seal (Mirounga) – but none of these
genera are known from the Pliocene marine mammal record there. Instead, we have
extinct phocids like Hadrokirus, Piscophoca, and Acrophoca;
these pinnipeds are known from both Peru
and Chile (with
the exception of Hadrokirus, which is so far only known from Peru).
Numerous other strange late Neogene marine mammals that are now extinct are
known from both Chile
and Peru,
including Brachydelphis (short snouted river dolphin - see above), Odobenocetops
(walrus faced whale), and Thalassocnus (aquatic sloth). These faunal
similarities attest to some sort of faunal turnover during the
Pliocene-Pleistocene interval. Valenzuela-Toro et al. suggested that phocids
were extirpated in this region as a result of sea level changes and changing
coastal geomorphology, with otariids repopulating the region as rocky
shorelines proliferated during the Pleistocene. I really enjoyed this paper,
and this was one of my first reviews for a paper in JVP; additionally, I had
the good fortune of meeting Anita in person at SVP in Los
Angeles this year; we spent some time looking at
pinniped fossils at the Cooper Center,
something I ought to write some blog posts about.


Miocene seagrass and sea cow fossil records.
The fossil record of seagrasses is relatively limited, and
little can be said directly from the fossil record of seagrasses regarding
their paleogeographic distribution. Seagrasses are the primary food source of
sirenians, and their modern distribution is tied to the distribution of
seagrasses. This new study by Jorge Velez-Juarbe uses the fossil record and
paleogeographic distribution of sea cows to reconstruct the paleogeography of
seagrasses through Cenozoic history. The sirenian fossil record suggests that
seagrasses were already distributed widely in the western North
Atlantic and Caribbean by the middle
Eocene. Oligocene cooling appears to have contracted the range of seagrasses
and sea cows, which soon expanded again early in the Miocene. Later in the
Miocene both groups expanded west and south into the eastern Pacific and the
western South Atlantic. The distribution of seagrasses –
as reconstructed from sea cow fossils – reached its modern pattern during the
Miocene.