Update 1, 2020: Back
in early 2014 I wrote this post with a much more limited body of available
literature - there were several things I knew about at the time but could not
discuss. So, six years later, after the publication of Llanocetus, Mystacodon,
Olympicetus, Fucaia, and others, it's time to revisit and update this post.
Update 2, 2024: Back in 2020 I updated this post, and some more discoveries have been made - including the recent work on the "Charleston Toothed mysticete" Coronodon, and the publication of Fucaia humilis - the oldest record of Neoceti from the North Pacific, and only Eocene mysticete from the northern hemisphere. I'm optimistic that I'll need to update this again!
Update 2.5, 2024: I realized I forgot to outline a possible Eocene dolphin vertebra reported by Davydenko et al., and have no included this.
Baleen whales (Mysticeti) and toothed whales (Odontoceti) are nearly
universally considered to share a sister-group relationship, and constitute a
monophyletic clade termed Neoceti (also known as Autoceta). Odontocetes and
mysticetes are generally considered (until recently) to have diverged and
diversified during the Oligocene, and neither group really has an extensive
fossil record prior to the Oligocene. On the other hand, archaeocetes – a
paraphyletic assemblage of stem cetaceans leading up to neocetes – generally
are considered to be restricted to the Eocene (though conference abstracts from
my doctoral lab by Josh Corrie and my adviser Ewan Fordyce suggest that
kekenodontids are late surviving Oligocene archaeocetes). For paleocetologists,
the Eo-Oligocene boundary is colloquially thought of as the archaeocete-neocete
split. However, many early Oligocene neocetes are relatively derived, and few
are really archaeocete-like – in other words, few appear to exhibit
archaeocete-like morphology with only a couple of acquired neocete
synapomorphies. This begs the question of when exactly Neoceti evolved.

The holotype skull of the early Oligocene dolphin Simocetus rayi - from the Alsea Formation of Oregon. I photographed the original specimen in spring 2016 at the Smithsonian.
Early
Oligocene Neoceti: Alsea Formation, Oregon
The majority of the Oligocene record of fossil cetaceans is limited to the late
Oligocene (owing to generally low sea levels and widespread erosion of early
Oligocene strata), but a few notable records of early Oligocene cetaceans are worth
discussing. In particular are the fossil cetaceans of the Alsea Formation in
west central Oregon. Although considered by Fordyce (2002) to be late
Oligocene, paleomagnetic data indicate it is probably early Oligocene in age
(Prothero, 2001). First and foremost is Simocetus rayi – the only
formally described cetacean from this unit, a bizarre agorophiid-grade dolphin
described by Fordyce (2002). Simocetus is pretty derived, and not really
archaeocete-like in many regards. Another agorophiid-grade dolphin is an
unnamed tusked odontocete, preliminarily reported by Fordyce et al. (2012) at
the Society of Vertebrate Paleontology meeting in Raleigh, North Carolina. This
undescribed dolphin is also fairly derived and fairly removed from the
odontocete stem. (Reidentified by Peredo et al. 2019 as a xenorophid, which I
don't necessarily buy - given that xenorophids are highly autapomorphic and
restricted to Oligocene strata in the Carolinas). A third cetacean from the
Alsea Formation is the world’s earliest toothless mysticete (an eomysticetid in
my personal opinion), recently named Maiabalaena nesbittae by Peredo et
al. (2018). A fourth cetacean which I spotted at the USNM in 2012 is an
undescribed aetiocetid with a complete braincase and somewhat basilosaurid-like
mandible. Yet another aetiocetid is known, but is currently under study by C-H
Tsai and myself, and I won’t spoil it. The Alsea Formation demonstrates that
the early Oligocene was populated by the same sorts of cetaceans known from the
late Oligocene. Most notably, the presence of an early chaeomysticete (Maiabalaena)
– in my opinion an eomysticetid – indicates that mysticete evolution is
telescoped into a 5 million year interval (or less), given our current
understanding of the timing of mysticete origins.

The two species of Fucaia from the Oligocene of Washington - not to scale! On the left is Fucaia buelli (UWBM collections) from the early Oligocene (Makah Formation), and on the right is Fucaia goedertorum (LACM collections), from the "middle" to late Oligocene Pysht Formation. Both specimens were discovered and collected by the Goederts.
The two original skulls of Olympicetus avitus published by Jorge Velez-Juarbe. Olympicetus is not really discussed in this post, but there are undescribed Simocetus/Olympicetus like dolphins from the older Makah Formation. Image by Jorge Velez-Juarbe.
Life reconstruction of a juvenile cf. Olympicetus specimen we
published - and which we hypothesized was unable to echolocate - and
therefore reconstructed here without a melon, and a bit of an
archaeocete like face (albeit much, much cuter!). Artwork by me!
A partial skill with associated mandible, vertebrae, and bulla of a Simocetus-like dolphin from the Makah Formation - from CCNHM collections. Collected, and generously donated to us, by Jim Goedert. This was my first serious acid preparation project - this is after a couple months of acid prep. It's been fully prepped out for a couple years and I'm working on describing it - it's one of the most plesiomorphic odontocetes in existence and has some very... interesting teeth.
Early
Oligocene Neoceti: Makah Formation, Washington
The lower
Oligocene Makah Formation is one of three Oligo-Miocene units on the strait of
Juan de Fuca on the north coast of the Olympic Peninsula of Washington; it is
overlain by the “middle” to upper Oligocene Pysht Formation and in turn by the
lower Miocene Clallam Formation. The Makah is about the same age as the Alsea,
but with a slightly older base at about 33.2 - 31 Ma. The first formally
described cetacean from the Makah Formation is the tiny toothed mysticete Fucaia
buelli named by Felix Marx et al. in 2015. It is quite similar to Chonecetus
goedertorum from the overlying Pysht Formation, which the authors
reassigned to Fucaia. These toothed mysticetes still had shearing wear
facets on their teeth, and may not have had baleen (as proposed for other
aetiocetids like Aetiocetus and Morawanacetus; though I will
cover this later).

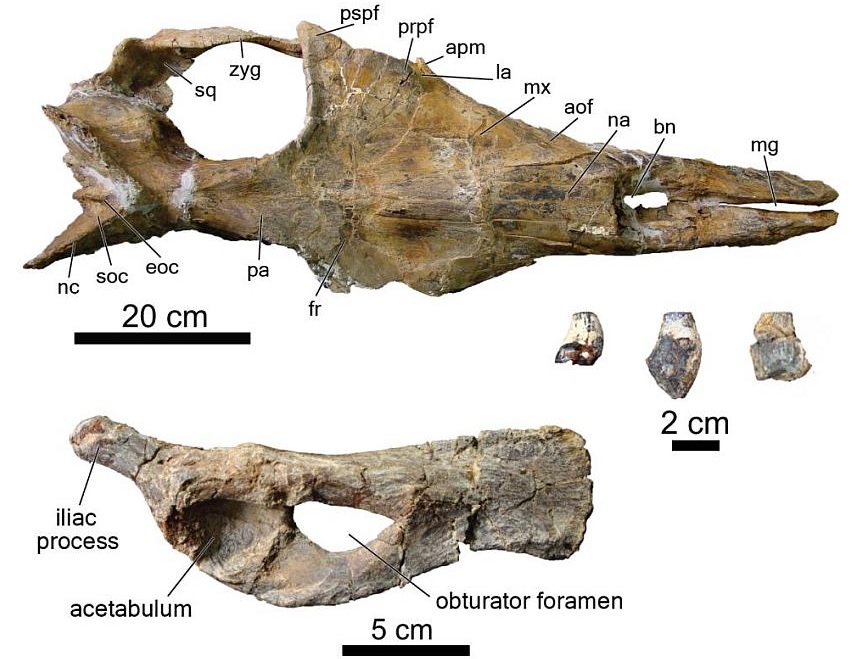
One of the most plesiomorphic (primitive) looking baleen whales ever discovered - Coronodon havenstein, from the lower Oligocene Ashley Formation. It's basically 75% archaeocete in terms of its skull shape, but we think it was able to filter feed with its teeth (no proto-baleen). On display at CCNHM. Mystacodon is slightly more primitive in its morphology but usually plots out a node further crownward (as a sister taxon to Llanocetus).
Also from the Ashley Formation - straight from the title of the paper - "an unexpectedly derived odontocete" - Ediscetus osbornei. This is a Waipatia-like or waipatiid-grade dolphin from the Ashley - at least a couple million years older than the oldest waipatiids from NZ, but possibly somewhat more derived owing to a more telescoped vertex. From Albright et al., 2018.
Early
Oligocene Neoceti: Ashley Formation, South Carolina
The Ashley
Formation is somewhat younger than the Alsea or Makah formations, but is known
from a substantially more diverse cetacean fauna of late early Oligocene age
(e.g. late Rupelian - ~30-28 Ma). Mysticetes include Coronodon havensteini, the
eomysticetid Micromysticetus rothauseni, and at least one strange
chaeomysticete I collected a couple years ago that may be related to the
fragmentary but strange New Zealand mysticetes Toipahautea and Whakakai.
Odontocetes include Agorophius pygmaeus, a handful of xenorophids (Xenorophus
sloanii, Albertocetus meffordorum, Inermorostrum xenops), the
recently re-named Ankylorhiza tiedemani, and one of the most archaic
known dolphins ever discovered – Ashleycetus planicapitis. Several
additional odontocetes, including the simocetid-like rostrum referred to Agorophius
pygmaeus by Fordyce (1981), and a dwarf Agorophius, are also known
and await description by yours truly. Most recently, and most surprisingly, is
a Waipatia-like dolphin, Ediscetus osbornei – possibly belonging
to the Waipatiidae, but there have been some problems with diagnosing and
defining this clade. Waipatia and Ankylorhiza
are the most derived odontocetes from the Ashley Formation – yet again, pushing
even more nodes and divergences further back into the early Oligocene. Perhaps what is most shocking about the Ashley Formation is its diversity: there are over a dozen species of cetaceans present. In contrast, the most diverse Eocene cetacean assemblages have half that (~5-6) - and most have only a quarter of that diversity (~3 species, like the Jackson Group on the Gulf coastal plain of the USA).
Early
Oligocene Neoceti: Ototara & Amuri imestones, New Zealand
An
interesting skull, the "protosqualodont of Keyes", was collected from
a famous limestone quarry near Oamaru (Parkside Quarry*) on the South Island of
New Zealand. I never ended up going to that quarry, since the limestone is
shockingly vertebrate poor - and I cannot recall if discovered from the quarry
or an exposure elsewhere of the Ototara Limestone, but the gigantic thick-boned
penguin Pachydyptes ponderosus was also discovered in this unit.
The "protosqualodont" of Keyes is a likely llanocetid, and one of the oldest known Neoceti from New Zealand. I cannot for the life of me find the published image of the skull in a block, so here are a couple of the preserved teeth of OU GS 10897 from Keyes (1973).
The
"protosqualodont" was originally reported by Ian Keyes (1973), who
was best known for publishing an excellent series of articles on fossil sharks
from New Zealand. The teeth were quite strange - at the time, a number of
poorly known isolated cheek teeth of highly variable shapes had been reported
from the Oligocene and lower Miocene of Australia and New Zealand, with many
named as different species of Squalodon, Parasqualodon, and Metasqualodon
- all tooth-based taxa. Mention these in front of Ewan and he'll furrow his
brow and say something like "quite difficult taxa, pity". Indeed,
most of these are isolated teeth and difficult to diagnose or even assess in
terms of diversity. These assemblages were all we had for decades in Australia
and New Zealand - and now appear to represent odontocetes, toothed mysticetes,
and kekenodontids (I will not be specific, owing to unpublished research). The
"protosqualodont" was different, though, because it was a set of
associated teeth. At the time, Keyes (1973) considered these the oldest
cetacean remains from New Zealand - indeed, the Ototara Limestone dates to
33-32 Ma, which is approximately earliest Oligocene. Originally discovered in
the matrix associated with a bone fragment - and misidentified as a partial
mandible by Keyes owing to its flatness, and later reidentified in Fordyce's
(1979) Ph.D. thesis as the top of the skull. The presumed molar is quite low
crowned, with very rugose enamel, and a vestigial third root; the top of the
braincase is generally similar to mammalodontids in its flatness. Later, Ewan
visited a private collector who happened to have a partial cetacean skull in a
block of limesstone from the same quarry - which happened to be the rest of the
skull! This specimen, OU GS 10897 - now appears to be a small bodied
llanocetid-like toothed mysticete.
*Parkside
Quarry is a source of bryozoan limestone facies of the Ototara Limestone -
known commercially locally as "Oamaru stone", which is sought after
for its desirable characteristics for stone carving. Many Māori artists use
the stone to carve koru (a spiral shape after unfurling tree fern fronds, a
symbol of new life/growth), wheku (stylized carving of a face), and Muri paraoa
(a whale tail; I received a greenstone carving of one from Ewan on my day in
NZ, a tradition for advisers to give to their students, and I have it hanging
on my PhD diploma in my office). I always wanted a Muri paraoa carving for
obvious reasons, but we could never afford one.
ZMT 67, a partial toothed mysticete mandible from the Amuri Limestone, South Island, New Zealand. This specimen is one of the oldest Neoceti from Australasia and is a possible llanocetid according to some of our analyses in Geisler et al. (2017) - photos from a book chapter by R.E. Fordyce.
Another
specimen from New Zealand is a mandible fragment from the earliest Oligocene
Amuri Limestone, which Fordyce (1989) identified as a possible toothed
mysticete. The specimen preserves some partial teeth, which had numerous cusps
in a radial pattern (a mysticete synapomorphy we proposed in our 2017 paper on Coronodon),
double rooted molars, and embrasure pits – the mandible is also very shallow,
unlike the tall, narrow mandible of Llanocetus. We actually coded the
highly fragmentary specimen into our cladistic analysis for Coronodon,
and it came out between Coronodon and aetiocetids (like Fucaia) - far removed from Llanocetus. However, our codings for Llanocetus were based on what was available from the literature before the skull was published upon - codings of the available mandible, and skull codings from Fitzgerald (2010). So, this position has changed. I personally think that ZMT 62 is probably a small bodied llanocetid.
In summary - while not as common as fossil assemblages dating to the late Oligocene, early Oligocene cetacean assemblages include at least a few species of mysticetes, an eomysticetid, and a few species of odontocetes. There are a few interesting things to consider: 1) early Oligocene faunas, despite being depauperate relative to late Oligocene faunas, still show some inkling of diversity - the late early Oligocene Ashley Formation being the most extreme (~10-15 cetaceans). 2) Many of these early Oligocene cetaceans are quite derived - examples include aetiocetids (Fucaia), eomysticetids (Maiabalaena, Micromysticetus), agorophiid-grade dolphins (Ankylorhiza), and waipatiid grade dolphins (Ediscetus). These taxa co-occur with more plesiomorphic cetaceans (e.g. Simocetus, Coronodon, Xenorophidae) and imply some ghost lineages at least a few million years long. 3) Another major contrast is body size - most of these, with the exception of Llanocetus, Coronodon, eomysticetids, and Ankylorhzia, are small. Most Oligocene odontocetes are smaller than a modern bottlenose dolphin; most toothed mysticetes, the mammalodontids and aetiocetids in particular, are downright tiny: harbor porpoise sized at the smallest (Fucaia buelli) and only a few that exceed a bottlenose dolphin in size (Aetiocetus), approaching the size of a pilot whale. We're still working this one out, but in concert with an explosive diversification in feeding morphology, it's likely related to changes in feeding and ecology across the Eocene-Oligocene boundary. 4) There is evidence of regional differences in faunas as early as the early Oligocene: simocetids and aetiocetids dominate North Pacific cetacean assemblages; xenorophids, agorophiid-grade dolphins, and Coronodon are only in North Atlantic assemblages; and New Zealand and Australia preserve mammalodontids, llanocetids, kekenodontids, and waipatiids (the latter shared with the North Atlantic). Only the Eomysticetidae are found in every basin during the Oligocene (now found in Oligocene rocks from Australia, New Zealand, Japan, Washington, Baja California (Mexico), North and South Carolina, and Austria. All of this evidence from the early Oligocene is highly suggestive of a many ghost lineages that should be illuminated by further collecting from earlier strata.

Holotype mandible fragment of the late Eocene
mysticete Llanocetus
denticrenatus from the La Meseta Formation of Seymour Island. From Mitchell (1989).
The large, and bizarre, skull of Llanocetus denticrenatus; latest Eocene, La Meseta Formation, Seymour Island, Antarctica. From Fordyce and Marx (2018).
Known
records of neocetes in the Eocene
As could
be surmised from the presence of relatively derived Neoceti in the early
Oligocene, a few – but only a few – records of latest Eocene mysticetes and
odontocetes exist. First and foremost, the only named Eocene neocete is the
earliest known baleen whale, Llanocetus denticrenatus from the La Meseta
Formation of Seymour Island in Antarctica. It was named by Ed Mitchell in 1989,
and the miserable scraps he designated as the holotype were originally
collected in the mid 1970’s. My Ph.D. adviser, Ewan Fordyce, returned to the
locality in 1986 and collected the rest of the specimen, which is a rather
large skull and mandible in addition to some postcrania. The bizarre skull,
complete with a markedly flattened rostrum, teeth with some basilosaurid-like
features and very wide diastmata (gaps between teeth), was recently published
by Fordyce and Marx (2018). The type specimen dates from just below the
Eo-Oligocene boundary and is approximately ~34 Ma in age. Accordingly, Llanocetus
is quite archaic - though nowhere near as basilosaurid-like as Coronodon
havensteini, which we named in 2017 (Geisler et al.); accordingly, Llanocetus
typically plots out a node or so crownward of Coronodon, despite being
older - suggesting that the lineage split even earlier during the Eocene. Shortly after the formal publication of the skull, more fragmentary material was repoted by Marx et al. (2019), including a tooth that is quite a bit larger than the same position in Llanocetus denticrenatus. This specimen, identified as Llanocetus sp., was collected by an Argentinean team, and hypothesized to represent a truly monstrous whale with a possible body length of 12 meters - exceeding the size of most archaeocetes, small extant baleen whales, all other toothed mysticetes, and approaching the size of Basilosaurus cetoides and some of the medium sized baleen whales around today.

An apparent second, and larger (but as yet, unnamed) species of Llanocetus exists - discovered recently and known mostly from isolated teeth. Interestingly, the new tooth bears a somewhat closer shape to that of the "protosqualodont" of Keyes (1973). The premolar of Llanocetus denticrenatus is on the left, and the premolar of the unnamed larger taxon, Llanocetus sp., is on the upper left.
In early
2017, the world was introduced to Mystacodon selenensis – a medium sized
toothed mysticete with a narrow rostrum from the uppermost Eocene Yumaque
Formation of Peru. Mystacodon has basilosaurid-like flippers, but with a
locked elbow joint – a clear neocete feature. The teeth are highly heterodont
and similar to a basilosaurid. The skull is decidedly basilosaurid-like but has
a slightly more telescoped vertex with a triangular occipital shield, and more
posteriorly placed nares - surprisingly similar to a skull of a NZ kekenodontid
(OU 22294) figured by Clementz et al. (2012). Subsequent studies have supported
a close relationship with Llanocetus, with Fordyce and Marx (2018)
assigning it to the Llanocetidae along with the “protosqualodont”.
The skull, teeth, and beautifully preserved pelvis of the basilosaurid-like toothed mysticete Mystacodon selenensis from the uppermost Eocene Yumaque Formation of Peru. From Muizon et al., 2017.
There is
also an undescribed odontocete preliminarily reported from about the
Eo-Oligocene boundary within the Lincoln Creek Formation in Washington state,
U.S.A. (Barnes et al., 2001). Another specimen, collected by Jim Goedert,
is a putative chaeomysticete from the late Eocene part of the Lincoln Creek
Formation – if the dating is accurate.* These specimens – although only barely
scraping into the Eocene – do demonstrate that odontocetes and mysticetes did
evolve before the end of the Eocene.
*Ironically,
I first learned of this specimen when I first wrote this post six years ago; I
received an email out of the blue from Jim Goedert. I then saw the specimen in
person at the Burke Museum in January 2016 – it’s definitely an
eomysticetid-like chaeomysticete, and has toothless mandibles and a poorly
telescoped braincase. This specimen is highly fragmented and needs lots of
careful preparation, and is begging for study. If accurately dated, it would
upend even much of the divergence dates pushed back by the discoveries outlined
here in this blog post. See “Caveats” for more.
A small vertebra (scale bar = 5 cm) from the uppermost Eocene La Meseta Formation of Seymour Island, Antarctica - likely assignable to Neoceti. But, could it be an early odontocete record? From Davydenko et al., 2020.
A latest Eocene odontocete from Seymour Island, Antarctica?
An additional possible neocete was recently reported by Davydenko et al. (2020), who reported an unusually small lumbar vertebra from the top of the La Meseta Formation of Seymour Island, Antarctica. This vertebra is small for a basilosaurid, though could be from a small basilosaurid like Zygorhiza, Dorudon, or Saghacetus - it has a slightly larger centrum, measuring about 8 cm wide and 6 cm long. However, and most critically, it differs from all basilosaurids, and indeed all known archaeocetes, in possessing a ventral keel. In basilosaurids, the ventral side is flattened to slightly convex. However, most published fossils of early Neoceti, along with many extant species, have a pronounced ventral keel on most or all of the lumbar vertebrae. When this paper came out, I thought this was a pretty profound revelation: I had not read this observation before, anywhere, and it's pretty easy to check for and, most critically, vertebrae are far more common than skulls. A few years later I decided to dive in a little further and used this character in our 2023 analyses of the phylogeny of mysticetes and odontocetes in the monographs on Coronodon and Xenorophus - and confirmed that it's a pretty useful synapomorphy of Neoceti, or a slightly more inclusive clade, depending upon which taxa fall outside Mysticeti (see below). Further evidence of Neocete affinities include the relatively short centrum, anteroposteriorly long transverse processes, a narrow triangular neural foramen, and well-developed and enlarged zygapophyses - more greatly resembling many Oligocene odontocetes and mysticetes than any basilosaurid. As tantalizing as this is, however, it's still a single vertebra hinting, or rather, teasing us at the prospect of finding some early odontocete fossils from Antarctica that could reveal the secrets of their early evolution.

The earliest known mysticete from the North Pacific, and the first such record from the Eocene - Fucaia humilis, discovered by my colleague Jim Goedert from uppermost Eocene strata of the Lincoln Creek Formation.
Most recently, my Ph.D. labmate Cheng-Hsiu Tsai invited me to collaborate on a study reporting a new species of the small aetiocetid Fucaia that had been collected by Jim Goedert from near the Eocene-Oligocene boundary within the Lincoln Creek Formation of southwestern Washington. The specimen was found as float on a river bank, though freshly exposed and not in a concretion that had traveled far downriver, and matching matrix with a slump of Lincoln Creek Formation. This particular horizon is well in the middle of Chron C13r based on prior paleomagnetic surveys, giving the specimen an age range of 35.1-33.7 Ma; the Eocene-Oligocene boundary is at 33.9 Ma, making the majority of this reversal within the Late Eocene (Priabonian stage). Based on the position, and assuming a continuous rate of deposition, we suggested an age of about 34.5 Ma. The appearance of toothed mysticetes is locally coincident with the oldest known fossil kelp holdfasts, suggesting a possible link between the earliest Neoceti of the North Pacific and the initial diversification of Fucalean kelps.
Are some toothed mysticetes actually misidentified archaeocetes?
One major, MAJOR caveat to all of this is the possibility that such beasts as Llanocetus, Coronodon, and Mystacodon might not actually belong within Neoceti. These fears were raised first by one of my other Otago PhD labmates, Josh Corrie, in his paper 2022 redescribing Kekenodon onamata with our Ph.D. adviser Ewan Fordyce. According to the phylogenetic analysis in this study, Kekenodon was recovered as an archaeocete, which is perhaps not surprising (especially when you consider the morphology of more completely preserved, but as yet undescribed kekenodontids that Josh is still working on). However, their analysis also recovered Llanocetus, Mystacodon, Coronodon, Janjucetus, and Mammalodon outside the clade Neoceti - rather than positioned within Mysticeti as recognized in earlier studies. What the hell? All of these have been well-documented mysticetes! How could we be so wrong? What the hell happened?
There are a few issues at stake here. First and foremost, these taxa are admittedly quite archaeocete-like - especially Mystacodon and Coronodon. So, there's only going to be a handful of features that pull them into the Neoceti - chiefly some features of the orbit region, earbones, and elbow. Second, the bootstrap support values for these basal nodes have always been weak - but have gotten weaker and weaker; in my own analyses, as I've coded more basal taxa into the matrix and more characters, the support values have eroded more and more. What that means is - clades that were once moderately well supported do not appear so any longer - suggesting that maybe these clades are not the answer after all. When support is that low, a few differences in codings, new characters, or the right new fossil can pull a bunch of lineages out.
Which brings us to the third problem: what is Neoceti? Strictly defined, it's the baleen whale-dolphin clade - Odontoceti + Mysticeti. All members of Neoceti, by definition, have to belong to one of these two groups. There's no middle ground; these Eocene-Oligocene fossils (broadly speaking) are either basal odontocetes, basal mysticetes, or they're archaeocetes. Perhaps we've made some wrong assumptions in the past about what's an archaeocete and what's a neocete. Many of these taxa have been known but under study for years - Coronodon and Llanocetus, and to a lesser extent, Mystacodon - are all prime examples of this. In the absence of including these in phylogenetic analyses (there were a couple studies, however) - many synapomorphies of the Neoceti were identified: presence of an antorbital notch, lack of exposure of the periotic on the lateral wall of the skull, locked elbow joint - among others. These features were based on observations of these early toothed mysticetes, along with early odontocetes like Simocetus and Waipatia. In other words, we knew what features defined the Neoceti. On the other hand, a consensus also emerged that Basilosauridae was the sister taxon to Neoceti. So, if it's not a basilosaurid, it has to be a neocete. Corrie and Fordyce (2022) suggested instead a third, somewhat radical hypothesis: that these taxa are also archaeocetes, just ones that are later diverging than the basilosaurids. After all, the possession of features thought to diagnose the Neoceti will be present in odontocetes and mysticetes - but could have evolved earlier - meaning that, depending upon the characters and taxa in a phylogenetic matrix, these taxa can pop in and out of Neoceti and onto the archaeocete "stem".

Character evidence supporting placement of Coronodon as a mysticete (black boxes, EW - equal weighting cladogram), as an archaeoicete (white boxes, IW - implied weighting cladogram) - or both (gray boxes). From Boessenecker et al. (2023).
We found some evidence of this when we published our huge monograph on Coronodon last year. Under equal weighting, these early taxa traditionally labeled as toothed mysticetes were placed within Mysticeti. However, under implied weighting - where the analysis software automatically down-weights problematic characters it reconstructs as being 'homoplastic' (e.g., sloppy, and evolving back and and forth many times) - Coronodon and friends (Llanocetus, Mystacodon) were recovered as archaeocetes. We reviewed the characters that drove this, and found that different suites of characters supported the Coronodon as a Mysticete v. Coronodon as an archaeocete hypotheses. Anatomical features supporting Coronodon and co. as mysticetes were generally features of the dorsal part of the skull and the basicranium, while features supporting placement outside of Neoceti were dominated by dental features. Many, if not most, of the features previously identified in Coronodon and other toothed mysticetes as synapomorphies of Neoceti were congruent with BOTH phylogenies!
So, here's the wrinkle: if this hypothesis is correct - and it might be - it would mean that virtually all Neoceti, with the exception of the recently named Fucaia humilis, are actually restricted to the Oligocene after all. What it does not change is the fact that the North Atlantic and Tethys are dominated by much more primitive basilosaurids and that possible/confirmed Neoceti are pretty much only known from the Pacific basin. No matter what we discover to be true in the long term, Llanocetus and Mystacodon are decidedly much closer to the Neoceti than basilosaurids are. To read more about this, check out last year's blog post about the phylogeny of Coronodon.
The
calibrated phylogenetic tree of Cetacea with molecular divergence dates
(horizontal gray bars) - showing the basamost nodes of Neoceti
diverging in the Eocene. A middle-of-the-road result, albeit based on
the time on fossils that had not been published. From McGowen et al.
(2009).
What
does Molecular divergence dating tell us?
Molecular
clock dating is a popular – if sometimes limited – method of estimating
divergences between clades. Unfortunately, we’re obviously limited to branches
between extant species – and in many cases, morphological phylogenies do not
recover the same relationships between extant species – calling into question
over which has it right. Another issue is that the rate can be very different
from what we see in the fossil record – with molecular divergence dates
sometimes pre-dating paleontological evidence by tens of millions of years (the
origin of modern placental mammals relative to the Cretaceous-Paleogene
extinction is a classic example). Unless properly calibrated with carefully
selected fossils with a clearly known phylogenetic position, divergence dates
can be off quite a bit.
Many
studies have inferred an Eocene divergence of Neoceti – and in some cases,
owing to the use of Llanocetus as a calibration point (e.g. McGowen et
al., 2009). Perhaps the most prominent cladistic analysis of cetaceans published
in recent years, Geisler et al. (2011) eliminated unpublished fossils from
their list of fossil calibrations – and in so were forced to ignore Llanocetus,
which at the time was only known by the original fragments published by
Mitchell (1989). The late Eocene age was clearly known – but relationships as a
toothed mysticete could not be resolved based upon the scrappy mandibular
fragment. As a result, the divergence of Neoceti was dated to the early
Oligocene. A recent analysis went so far as to date the divergence of crown
Odontoceti as occurring during the late middle Eocene – despite no crown
odontocetes being demonstrably known from the late Oligocene or earlier
(McGowen et al., 2020). In contrast, the more conservative analysis of Geisler
et al. (2011) recovers the divergence of Crown Odontoceti in the early
Oligocene. This is still interesting to me, as the earliest diverging extant
odontocetes – the sperm whales – have a strictly post-Oligocene fossil record,
with the possible exception of the fragmentary Ferecetotherium, known
mostly from teeth and a mandible from the latest Oligocene of Azerbaijan.
There are a *ton* of different studies I have glossed over, and there's likely something I've missed. Regardless,
at this point there is clear support from molecular clock analyses for an
Eocene origin of odontocetes. But how early?
The
earliest Basilosauridae – middle Eocene
One
problem inherent with a late or even latest Eocene origin of Neoceti is that it
would telescope the majority of basilosaurid evolution into a 5 Ma period
during the Priabonian and late Bartonian stages of the Eocene. The oldest
records of traditionally identified basilosaurids are only about 40 Ma, only
5-6 Ma older than Llanocetus. However, a recent discovery of a
basilosaurid from the Bartonian (late Middle Eocene) of Ukraine suggests a
reinterpretation of “Eocetus” wardii from similarly aged strata
in the eastern USA. Gol’din and Zvonok (2013) named a new genus and species, Basilotritus
uheni – which has vertebrae like “Eocetus” wardii with the
tympanic bulla of a basilosaurid. Gol’Din and Zvonok (2013) transferred “Eocetus”
wardii to Basilotritus, recombining it as Basilotritus wardii.
In addition to the recognition of both species of Basilotritus as early
basilosaurids, a couple other middle Eocene basilosaurids have been reported,
including a braincase from the Bartonian of New Zealand identified as Zygorhiza
sp. (Kohler and Fordyce, 1997), and Ocucajea and Supaycetus from
the Bartonian of Peru (Uhen et al., 2011). Unfortunately, the
protocetid-basilosaurid transition is poorly known, although several
protocetids exhibit derived basilosaurid-like features, including Georgiacetus,
Babiacetus, and Eocetus, and the earlier basilosaurids like Basilotritus
and Supaycetus are a bit more plesiomorphic than other
basilosaurids.

Phylogenetic relationships and stratigraphic
ranges of Basilosauridae, from Gol'Din and Zvonok (2013).
Along with
an earlier divergence of Neoceti, and the extremely primitive anatomy of early
toothed mysticetes like Coronodon and Mystacodon, the possibility is raised –
how different would the earliest neocetes actually look from basilosaurids? Are
basilosaurids actually before the split? The limited descriptions available for
some, and limited cranial material for most, could allow for this sort of situation.
Godfrey et al. (2012) go so far as to highlight that, in a description of a
partial skull of unknown age, that the earliest mysticetes and odontocetes
could even grossly resemble protocetids in their skull anatomy. This again is
the same problem highlighted above regarding molecular versus morphological
divergence: anatomical features may change much more slowly and manifest after
the molecular split, in which case we would have no way of knowing (save for
broader taxon sampling and extremely careful coding) if some basilosaurids were
in actuality stem odontocetes or stem mysticetes. Personally, all archaeocete
groups are admittedly paraphyletic anyway, and I see this as probable but
likely (at this point) intractable (but, see above).
Why are
there no late Eocene Neoceti from Egypt or the southeastern USA? Some thoughts,
and caveats
This is
the question that really chaps my ass. Where are the dolphins and mysticetes
from the Jackson Group of the gulf coast - latest Eocene - very well-sampled
strata - or, from the most meticulously sampled (and beautifully exposed)
whale-bearing late Eocene rocks anywhere in the world: Egypt?
The short
answer is, I have no clue. Long, rambling answer: It’s worth noting that late
middle and early late Eocene assemblages from South Carolina and Morocco have
some similarities (pappocetine protocetids, Chrysocetus), and there are
genus-level similarities with the late Eocene of the southeast USA and Egypt in
general (Basilosaurus, Dorudon, ?Cynthiacetus). It’s not
perfect – not like the similarities between Pliocene marine mammals from Europe
and the eastern USA – and perhaps archaeocetes were less pelagic than modern
cetaceans (hardly an unreasonable hypothesis), therefore having more regional
provincialism. With only a little data, it’s tempting to suggest that perhaps
the North Atlantic and Tethys were a single biogeographic province, dominated
by archaeocetes. These rocks, after all, are very tightly dated to the latest Eocene - there is abundant evidence that the Yazoo Clay of the Gulf Coastal Plain and the Qasr el Sagha Formation at Wadi al Hitan marks the last million or so years of the Eocene, and erosional surfaces within and at the top of the unit mark the Antarctic glaciation at the end of the Eocene.

What’s the
point of all this? All examples of Eocene Neoceti are either from the eastern
Pacific or from the Antarctic peninsula, on the fringes of the Pacific: Lincoln
Creek Formation of Washington, USA; Yumaque Formation, Peru; La Meseta
Formation, Seymour Island, Antarctica). Some bona fide archaeocetes are known
from late Eocene assemblages from the Pacific (Zygorhiza sp., New
Zealand; Cynthiacetus peruvianus, Peru), but these assemblages are not
exclusively dominated by archaeocetes like known assemblages in the
Atlantic-Tethyan province is. So, maybe there is a regional difference. In
which case – more Eocene rocks along the Pacific margin clearly need to be
explored. There are some Eocene marine deposits in California and Oregon [stay
tuned…] which are sure to eventually produce some cetacean remains. I’m also
convinced that more Eocene-Oligocene deposits probably exist further north in
the Pacific Northwest, in British Columbia and southern Alaska. There are tons
of Oligocene cetaceans from Japan, and no shortage of Eocene mollusks. I have a
hard time believing there were no cetaceans there during the Eocene. I also
wonder about the Korean peninsula and coast of China. On the other hand, where
there’s smoke, there’s fire: sometimes it’s easier to attack a known fossil
site then to discover new ones. In which case, further field investigation of
the Lincoln Creek Formation (and the Yumaque Formation of Peru) seems like an
obvious choice.
Lastly,
caveats. One issue that comes up time and again is that most of the ages for
deposits in the Pacific northwest stem from a single edited volume (Prothero,
2001) and I’ve heard many private grumblings about the quality of the
paleomagnetic analyses published within (politely refraining from repeating
them here), and read some published responses (Nesbitt et al., 2010). So, it’s
possible that these paleomagnetic analyses could be off by one or more reversed
or normal chrons – paleomagnetism is out of necessity done in concert with
biostratigraphy, typically foraminifera or perhaps diatoms. In the Pacific
Northwest, most of the micropaleontological work has been done with benthic
foraminifera rather than planktonic foraminifera – the latter is much, much
more accurate than the former, and so the biochronology of mid Cenozoic Pacific
Northwest stratigraphy is, for the time being, a bit hamstrung. So many of
these surprisingly derived simocetids and eomysticetids from the earliest
Oligocene may be a bit younger. That being said, this has no bearing on
cetaceans of similar evolutionary stage showing up in the 28-30 Ma Ashley
Formation, dated to late Rupelian with forams, strontium, and – if I recall
correctly – dinoflagellates as well. So, there would have to be *pervasive*
mis-dating of early Oligocene cetacean assemblages worldwide in order for
dating to be a viable problem.
Summary
In
summary, there are several derived neocetes (Ediscetus, Ankylorhiza, Maiabalaena, Micromysticetus) from the early Oligocene, and in some cases forming diverse assemblages, a couple
of genuine records of Neoceti from the latest Eocene (Mystacodon, Llanocetus)
– and lastly, the expanded fossil record of basilosaurids now ameliorates the
problem of a formerly telescoped record of the family. More records of early
odontocetes and mysticetes from the Eocene does not sound like such a strange
idea anymore, but is in fact now predicted by the fossil record. We have little
evidence of it, but improved sampling of late Eocene marine rocks – especially
from poorly sampled areas (in terms of Eocene rocks) like the Pacific Northwest,
the west coast of South America, and possibly Japan, the Korean peninsula, and
China – may yield more records of early Neoceti.
References
Cited
Albright
et al. 2018. https://www.tandfonline.com/doi/abs/10.1080/02724634.2018.1482555?fbclid=IwAR3hdOiPhWZ3haClIdaCjTF63DNWbM93u4zBiql8VrmXgGE-e63zxcplxBg&journalCode=ujvp20
L. G.
Barnes and J. L. Goedert. 2000. The world's oldest known odontocete (Mammalia,
Cetacea). Journal of Vertebrate Paleontology 20(3):28A.
Boessenecker, R.W., B.K. Beatty, and J.H. Geisler. 2023. New specimens and species of the Oligocene toothed baleen whale Coronodon from South Carolina and the origin of Neoceti. PeerJ 11:e1479. https://peerj.com/articles/14795/
Davydenko, S., Mors, T., and P. Gol'din. 2020. A small whale reveals diversity of Eocene cetacean fauna of Antarctica. Antarctic Science. https://www.cambridge.org/core/journals/antarctic-science/article/small-whale-reveals-diversity-of-the-eocene-cetacean-fauna-of-antarctica/60762DFF372D829534013A54D4BEF066
Fordyce, 1989. https://www.tandfonline.com/doi/abs/10.1080/00288306.1989.10425719
R. E.
Fordyce. 2002. Simocetus rayi (Odontoceti, Simocetidae, new family); a
bizarre new archaic Oligocene dolphin from the eastern North Pacific. Smithsonian
Contributions to Paleobiology 93:185-222.
Fordyce
and Marx, 2018. https://www.sciencedirect.com/science/article/pii/S096098221830455X
R.E.
Fordyce, E.M.G. Fitzgerald, G. Gonzalez Barba. 2012. Long-tusked archaic
odontocetes from Oregon and Baja California Sur, eastern Pacific Margin.
Journal of Vertebrate Paleontology 32:3:95.
Geisler et al., 2011. https://bmcevolbiol.biomedcentral.com/articles/10.1186/1471-2148-11-112
Geisler et
al., 2017. https://www.sciencedirect.com/science/article/pii/S0960982217307042
Godfrey et al., 2012. https://anatomypubs.onlinelibrary.wiley.com/doi/full/10.1002/ar.22637
P. Gol'din
and E. Zvonok. 2013. Basilotritus uheni, a New Cetacean (Cetacea,
Basilosauridae) from the Late Middle Eocene of Eastern Europe. Journal of
Paleontology 87(2):254-268.
Keyes, 1973. https://www.tandfonline.com/doi/abs/10.1080/00288330.1973.9515482
R. Kohler
and R. E. Fordyce. 1997. An archaeocete whale (Cetacea: Archaeoceti) from the
Eocene Waihao Greensand, New Zealand. Journal of Vertebrate Paleontology
17(3):574-583.
Lambert et al., 2017. https://www.cell.com/current-biology/comments/S0960-9822(17)30435-9
Marx et al., 2015. https://royalsocietypublishing.org/doi/10.1098/rsos.150476
Marx et al., 2019. https://www.cambridge.org/core/journals/antarctic-science/article/gigantic-mysticete-predators-roamed-the-eocene-southern-ocean/0EEFC32753A8909BC4E7C134F5AEA6AE/core-reader
McGowen et al., 2009. https://www.sciencedirect.com/science/article/abs/pii/S1055790309003431?via%3Dihub
McGowen et al., 2020. https://academic.oup.com/sysbio/article/69/3/479/5601630
E. D.
Mitchell. 1989. A new cetacean from the late Eocene La Meseta Formation,
Seymour Island, Antarctic Peninsula. Canadian Journal of Fisheries and Aquatic
Sciences 46(12):2219-2235.
Muizon et
al., 2018. http://sciencepress.mnhn.fr/en/periodiques/geodiversitas/41/11
Nesbitt et
al., 2010. http://webcache.googleusercontent.com/search?q=cache:091QUEdyObUJ:ftp://ww4.dnr.wa.gov/geology/polenz/refs/Nesbitt%2520et%2520al%25202010_blakeley%2520Fm%2520age_Olympics%2520%2B%2520Vancouver%2520Isl_6-22-10.pdf+&cd=2&hl=en&ct=clnk&gl=us&client=firefox-b-1-d
Peredo et
al., 2018. https://www.sciencedirect.com/science/article/pii/S0960982218314143
Peredo et
al., 2019. https://www.cambridge.org/core/journals/paleobiology/article/convergence-on-dental-simplification-in-the-evolution-of-whales/29E0C07483DCB9108D18F96BE57A474B
Prothero
DR, Bitboul CZ, Moore GW, Niem AR. 2001. Magnetic stratigraphy and tectonic
rotation of the Oligocene Alsea, Yaquina, and Nye formations, Lincoln County, Oregon.
In: Prothero DR, ed. Magnetic stratigraphy of the Pacific coast Cenozoic.
Pacific Section SEPM (Society for Sedimentary Geology) 91:184–194.
Tsai, C.-H., J.L. Goedert, and R.W. Boessenecker. 2024. The oldest mysticete in the Northern Hemisphere. Current Biology 34:1794-1800. https://www.cell.com/current-biology/fulltext/S0960-9822(24)00308-7?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0960982224003087%3Fshowall%3Dtrue
M. D. Uhen
(2007): The earliest toothless mysticete: A chaeomysticetan
from the early Oligocene Alsea Formation, Toledo, Oregon. – Journal of Vertebrate
Paleontology 27/3 Suppl.: 161A.
M. D.
Uhen, N. D. Pyenson, T. J. DeVries, M. Urbina, and P. R. Renne. 2011. New
middle Eocene whales from the Pisco Basin of Peru. Journal of Paleontology
85(5):955-969.