
2023 has been the worst year of my adult life. Within the span of a month, we lost our cat Burt Reynolds to a stroke (we got him in New Zealand), we lost my father-in-law to brain cancer, and the work environment we had formerly enjoyed had deteriorated so much that it reached a tipping point and we both resigned from CofC. Later this fall, my Ph.D. adviser, Dr. R. Ewan Fordyce, passed away - robbed of a long, peaceful, and productive retirement. This year has really been a crucible - and what has emerged for us is a complete lack of tolerance for mismanagement, wage theft, academic bullying, and other issues we've faced in the past. This whole experience has been an exercise in taking punishment and setting boundaries. It hasn't been all bad: I got two enormous monographs published this year - one on the toothed mysticete Coronodon and the other on the early dolphin Xenorophus, both from the Oligocene of South Carolina.
Aside from those studies, this year has seen a number of great papers - another monograph on stem odontocetes by Jorge Velez-Juarbe on Washingtonian simocetid dolphins. Two new spectacular basilosaurids were reported: the tiny Tutcetus, and the absolutely monstrous Perucetus colossus - the latter with body mass estimates overlapping with and exceeding the blue whale. Our Ukrainian colleagues have managed to resume research life now that the situation over there has improved quite a bit - Svitozar Davydenko has succeeded in publishing three papers on archaeocete limbs this year. Calvert Cliffs volume II came out this year, five years after volume I, and includes a monograph reporting all of the fossil odontocetes known from the cliffs. Amber Coste has published two long-awaited papers on 'waipatiid' grade dolphins from the Oligocene of New Zealand from her thesis research - these new tusked dolphins, Nihohae and Nihoroa, are welcome additions to our pantheon of Oligocene whales. It was a slow year for pinnipeds and sirenians, unfortunately (but don't fret: there are some great papers that will be coming out next year!).
Now: I have possibly, if not probably, missed something. If I have, please let me know in the comments! But!: please check the 2022 review to make sure it was not a paper that came out in 2022 with 'online early' status and formally published this year. The usual disclaimers apply: I absolutely will not update the posts to shift a 2022 online early publication to this year; multiple papers by the same author are not necessarily in alphabetical order and you just have to accept that; lastly, the study must report fossil (or subfossil) material. Papers by paleontologists reporting modern osteological specimens are not paleontological studies and therefore do not make the cut. Also: I have not, and will not, type all of these out into journal reference format. And yes: I've been asked to do all of these things. Papers I formally reviewed are marked with an asterisk (a surprising minority this year!).

[Abstract in Japanese] 2022年6月5日,岐阜県瑞浪市釜戸町の土岐川河床に 分布する中新統瑞浪層群宿洞層からパレオパラドキシア 科(束柱目)の骨格化石が発見された(図1).骨格は少 なくとも腰椎,仙椎,右後肢が関節した状態で保存され,さらに一連の左肋骨遠位端の整然とした露出状況や頭部 も非常に近い位置に保存されていたことから胸部~頭部 の骨格も関節状態で保存されていることが予測され,さ らに河床に露出しているため標本流出の恐れがあったこ とから,6月10日に緊急発掘を実施した(図2). 現在,瑞浪市役所・教育委員会の関係課と研究者が連
携し,あわせて市内外の方々から様々な支援を受けなが ら「瑞浪産パレオパラドキシア全身骨格化石復元プロジェ クト」を実施し,瑞浪市化石博物館で標本の剖出を進め ている.その結果,前肢を除くほぼ全身の骨格が関節 した状態を残して保存されていること,パレオパラドキ シア科に特徴的な円柱を束ねたような歯冠の低い臼歯が 上下顎に植立した状態で保存されていることが判明した (図3, 4).今後,骨格の復元,系統分類学的な位置づけ,古生態の復元,埋没過程の解明,古環境の解明など多岐 の分野にわたる研究を進めていく予定である.

Antar et al.: A diminutive new basilosaurid whale reveals the trajectory of the cetacean life histories during the Eocene.
Soon after whales originated from small terrestrial artiodactyl ancestors, basal stem forms (archaeocetes) came to inhabit more specialized aquatic ecologies and underwent a tremendous adaptive radiation that culminated in the adoption of a fully aquatic lifestyle. This adaptive strategy is first documented by the geographically widespread extinct family Basilosauridae. Here we report a new basilosaurid genus and species, Tutcetus rayanensis, from the middle Eocene of Fayum, Egypt. This new whale is not only the smallest known basilosaurid, but it is also one of the oldest records of this family from Africa. Tutcetus allows us to further test hypotheses regarding basilosaurids’ early success in the aquatic ecosystem, which lasted into the latest Eocene, and their ability to outcompete amphibious stem whales and opportunistically adapt to new niches after they completely severed their ties to the land. Tutcetus also significantly expands the size range of the basilosaurids and reveals new details about their life histories, phylogeny, and paleobiogeography.
Comments: A great new paper by my Egyptian colleagues, many of whom were coauthors on our earlier Phiomicetus paper led by Abdullah Gohar. The specimen itself is surprisingly small - scarcely larger than a bottlenose dolphin - and old, one of the older basilosaurid specimens. This alone is interesting as it overlaps in size with earlier protocetids. However, what I find most interesting about this new work is the discussion on ontogeny and dental eruption - Tutcetus - yes, named after King Tut - has permanent teeth that erupt relatively early in growth and suggest a precocial lifestyle. I see papers like this that delve into life history as a welcome expansion of topics we should be discussing in marine mammal paleontology (and they're topics I think about a lot).

In this article we describe a first find of a seacow, Halitherium sp. A marine mammal best known from Oligocene deposits from the Mainz Basin, Germany, among others. It concerns the proximal portion of a right upper arm bone, a humerus, heavily fossilized and graybrown in color, which was trawled from the estuary of the Western Scheldt, Zeeland, the Netherlands. We know that it was trawled by fishermen from the mouth of the Western Scheldt, but that area is quite large and includes, among others, the fossil sites of ‘De Wielingen’ (the Netherlands) and, as an extension of that navigational route, ‘Het Scheur’ (Belgium). Based on morphological differences and size, this proximal humerus can be easily distinguished from other Sirenia. In this article we also give an overview of the different members of the seacows (Sirenia) and their evolution.

The North Pacific rim was home to an extinct group of semiaquatic marine mammals, the order Desmostylia, which superficially resembled hippos. Desmostylians are an uncommon fossil vertebrate in most localities where they occur, and Oligocene taxa particularly so. Beyond the type dentition and two femora of Cornwallius sookensis, and the skull and postcrania of Behemotops proteus, both previously described from the region of Muir Creek locality of the upper Oligocene Sooke Formation of southern coast of Vancouver Island, British Columbia, Canada, no other records from this site have been reported. Additional specimens of teeth have now been found in the Royal British Columbia Museum and Canadian Museum of Nature, including a long-forgotten and unreported paratype. In total, three additional teeth, one of B. proteus and two possibly belonging to C. sookensis, are reported here from its type locality. More importantly, the upper molar reported here, as well as UCMP 36078 from Baja California and USNM 181740 from Oregon, all resemble each other but should be considered as belonging to Cornwallius only with caution.

Fossil cetaceans are often found in Miocene marine outcrops across the globe. However, because this record is not homogeneous, the dissimilar increase in occurrences, along with the sampling bias has created regions with extensive records and others with great scarcity. Among these, the Caribbean has remained enigmatic due to the lack of well-preserved cetacean fossils. Here, we report new Caribbean fossil cetaceans from the Upper Miocene Chagres Formation exposed along Piña beach, Eastern Panama, including a scaphokogiine kogiid, an Acrophyseter-like physeteroid and the phocoenid Piscolithax. Along with previous records of the iniid Isthminia panamensis and the kogiine Nanokogia isthmia, the Chagres cetacean fauna shows some similarities with other Late Miocene cetacean communities such as the Californias in the North Pacific, although their closest affinities lie with the eastern South Pacific Pisco Formation, Peru. Such findings indicate that though deep and intermediate Caribbean–Pacific water interchange was reduced by the Middle Miocene due to the shallowing of the Central American Seaway, shallow waters marine connection that persisted until the Pliocene might have facilitated the dispersal of coastal species across both sides of the Isthmus.
Comments: Not exactly on California fossils, but still an extremely welcome addition to eastern Pacific whaleontology: this study helps fill in a bit of a geographic gap during the late Miocene. Some of these taxa are known from the upper Miocene of California, like Piscolithax - and we find teeth of an Acrophyseter-like sperm whale from these deposits as well. Back in 2013 I compared and contrasted Pliocene/late Miocene marine mammal assemblages from around the world, and it's nice to see similar studies chasing that up (and getting into high profile journals, for that matter).

Bianucci et al.: A heavyweight early whale pushes the boundaries of vertebrate morphology.
The fossil record of cetaceans documents how terrestrial animals acquired extreme adaptations and transitioned to a fully aquatic lifestyle1,2. In whales, this is associated with a substantial increase in maximum body size. Although an elongate body was acquired early in cetacean evolution3, the maximum body mass of baleen whales reflects a recent diversification that culminated in the blue whale4. More generally, hitherto known gigantism among aquatic tetrapods evolved within pelagic, active swimmers. Here we describe Perucetus colossus—a basilosaurid whale from the middle Eocene epoch of Peru. It displays, to our knowledge, the highest degree of bone mass increase known to date, an adaptation associated with shallow diving5. The estimated skeletal mass of P. colossus exceeds that of any known mammal or aquatic vertebrate. We show that the bone structure specializations of aquatic mammals are reflected in the scaling of skeletal fraction (skeletal mass versus whole-body mass) across the entire disparity of amniotes. We use the skeletal fraction to estimate the body mass of P. colossus, which proves to be a contender for the title of heaviest animal on record. Cetacean peak body mass had already been reached around 30 million years before previously assumed, in a coastal context in which primary productivity was particularly high.
Comments: I was asked to comment on this by several journalists the day I tendered my resignation from CofC - my first day as a 'free agent' I spent traveling to spend time with my family at Lake Tahoe and speaking on the phone with journalists during my layover at O'Hare. Needless to say this paper has left an indelible imprint on me. It's become somewhat controversial and I don't really understand why: Perucetus is a colossal basilosaurid whale, and while it's not terribly complete, I think the estimates and methods used are appropriate. Most of the folks complaining about this study are dino-weenies who are probably just mad that yet another whale has outranked dinosaurs as the largest/heaviest animals ever.
Now, on a scientific level: just like Tutcetus, this gives a surprising expansion to the range of body forms in basilosaurid whales. Perucetus is as large or possibly even larger than Basilosaurus cetoides, and given how incredibly inflated and dense/heavy the bones are, it almost certainly had a very different ecology than other basilosaurid whales. The authors suggest three possibilities: shallow water benthic foraging on seagrasses or algae like sirenians, foraging on marine invertebrates, and scavenging on carcasses. The authors rightly indicate that herbivory is quite unlikely: the last prior herbivorous "cetacean" was Indohyus and such an ecology is otherwise unknown in cetaceans. The latter I think is unlikely - but admittedly quite similar to the T. rex as a scavenger hypothesis by Jack Horner. As a walrus expert, I am sympathetic to the second hypothesis - but all of these will require further discoveries from Peru to be made and most importantly, a skull with teeth.

The evolutionary history of the beaked whales (Ziphiidae), odontocetes nowadays adapted to deep diving, is well known thanks to a significant fossil record mainly from the deep ocean floors. A partial cranium of a ziphiid recovered from Plio-Pleistocene deep sea deposits (about 1000 m) off the port of Pisagua, northern Chile, during fishing activity is here described and referred to the new species Ihlengesi changoensis. Ihlengesi changoensis differs from the type species Ihlengesi saldanhae, from the sea floor off South Africa, by having a more elongated premaxillary sac fossa and consequently a more anteriorly located premaxillary foramen; dorsal margin of each premaxillary crest sloping markedly ventrolaterally and generating an acute dorsal profile of the vertex in anterior view; less anterolateral extension of the right nasal forming part of the premaxillary crest; lateral margins of the nasals not anteriorly diverging but weakly convex; nasofrontal suture anteriorly pointed. The phylogeny supports a sister-taxon relationship between I. changoensis and I. saldanhae, both members of the crown ziphiids Hyperoodontinae. Ihlengesi changoensis shares with I. saldanhae and other fossil ziphiids a small body size (estimated length 3.5 m) supporting the hypothesis that in the past small beaked whales (<4 m) were more common than today. Such recent shift of the ziphiids towards a larger size could be the result of a progressive change of diet from fish to cephalopods, to the competition with the delphinids, and the predatory impact of the white shark Carcharodon carcharias and/or of the killer whale Orcinus orca. This new Chilean ziphiid further supports the hypothesis that crown beaked whales originated and firstly dispersed in the oceanic waters of the Southern Hemisphere. Their radiation and geographical distribution could have been driven by the southern oceanic circulation and related localized concentration of trophic resources in high productivity upwelling areas.

A comprehensive revision of the localities of discovery of fossil mysticetes is presented together with a highly inclusive phylogenetic
analysis in order to provide the basis for a chronology of the main
mysticete diversification events. The results suggest that the origin of
Mysticeti (that include earlier toothed taxa together with today
baleen-bearing cetaceans) occurred c. 38 Ma; the origin of Chaeomysticeti (that include only baleen-bearing cetaceans) occurred c. 28 Ma; the origin of Balaenomorpha (crown mysticetes) occurred c.
23.3 Ma. Additional chronological inferences are provided. Within this
chronological framework, we analyzed diversity trends, origination and
extinction patterns and body size evolution, and looked for eventual
causal relationships between evolutionary processes, marine and
terrestrial ecological turnovers and geodynamic
events. We found four main diversification events corresponding to
peaks in originations and, in a few cases, with the origin of different
feeding strategies adopted by the differen mysticete families. We found
that different mechanisms are correlated to specific diversification
events and these include changes in temperature and ocean circulation
patterns, nutrient availability in the water column and diatom abundance
and diversity. Resuming, no single mechanism explains all the
diversification events occurred during mysticete evolution; rather, we
found that each diversification event was correlated to different
combinations of biotic and abiotic factors, suggesting that this group experienced major adaptation processes to the changing paleoenvironments in the last 38 Ma.

A partial skeleton of a Pliocene balaenid whale (Mammalia, Cetacea, Mysticeti) is described and compared to a large set of extant and fossil Balaenidae. The specimen (MCRE 232834) includes a jugal, both mandibular rami and part of the postcranial skeleton including several vertebrae, complete ribs, hyoid, pelvis, a single scapula and a single partial forelimb. The specimen was found at a site in the vicinity of the San Valentino Castle, about 16 km S from Reggio Emilia, close to the town of Castellarano, Emilia Romagna (northern Italy). Molluscs and foraminifers indicate a late Zanclean age for MCRE 232834, constrained between 3.8 and 3.6 Ma. A taphonomic analysis revealed that after death the individual sunk on the sea floor upside down and underwent a series of biostratinomic processes eventually leading to the collapse of the ribcage and to the disarticulation of the posterior thoracic, lumbar and caudal vertebrae, together with the loss of several skeletal elements including the skull. Shark teeth and encrusting molluscs demonstrate that the specimen was exploited by different organisms during its decay. The study of the skeleton revealed that MCRE 232834 shows an abruptly converging anterior ends of the mandibular rami, well-developed olecranon process in the ulna, peculiar morphology of the cervical vertebrae and enlarged attachment sites for axial muscles in the ribs. Based on the morphology of the cervical vertebrae, mandible and scapula, MCRE 232834 can be assigned to a new genus and species of the family Balaenidae, i.e., Charadrobalaena valentinae n. gen., n. sp., which is part of a primitive clade of balaenids that is the sister group of the crown balaenid whales. A functional analysis of the vertebral column revealed that it was able of comparatively faster and more agile swimming with respect to the extant balaenid species.

Boessenecker et al.: New specimens and species of the Oligocene toothed baleen whale Coronodon from South Carolina and the origin of Neoceti.
Baleen whales (Mysticeti) are gigantic filter-feeding cetaceans possessing the unique soft tissue structure baleen and lacking adult teeth; Oligocene fossils have revealed a wealth of early diverging tooth-bearing mysticetes highlighting the transition from archaeocete ancestors to early toothless baleen-bearing eomysticetid whales. The archaeocete-like, toothed mysticete Coronodon havensteini from the lower Oligocene Ashley Formation of South Carolina possesses a number of peculiar aspects of feeding morphology suggesting dental filter-feeding in the earliest diverging mysticete lineage. New fossils of Coronodon are described in detail, including (1) supplementary description of the holotype skull and skeleton of Coronodon havensteini; (2) description of two new juvenile skulls of C. havensteini and a partial skull and postcranial skeleton of an adult; (3) description of the new species Coronodon planifrons n.sp.; and (4) description of the new species Coronodon newtonorum. New specimens of Coronodon havensteini include a partial adult skeleton preserving new elements for the species including incisors, numerous upper premolars and molars, lower m4, scapula, lumbar, and caudal vertebrae, and two juvenile skulls with tympanoperiotics and teeth. Fossils from the overlying unit, the Chandler Bridge Formation, represent two new species: Coronodon newtonorum n. sp. and Coronodon planifrons n. sp. Coronodon newtonorum possesses a concave-up alveolar profile, a mandibular condyle elevated far above the toothrow, and a gracile periotic resembling those of juvenile C. havensteini. Coronodon planifrons n. sp. possesses a horizontal supraorbital process, successively smaller upper molars, massively inflated periotic, and longer intertemporal region. Coronodon planifrons n. sp. preserves one of the most complete vertebral columns among toothed mysticetes, indicating nine thoracic vertebrae, ten lumbar vertebrae, and at least 20 caudal vertebrae. The column exhibits a somewhat stabilized caudal peduncle with enlarged lumbocaudal vertebrae, and rectangular terminal caudals indicate the presence of tail flukes. Juvenile skulls reveal several ontogenetic trends in Coronodon havensteini, including the anterior migration of the orbitotemporal crest, anteroposterior elongation of the intertemporal region, inflation of the body of the periotic, enlargement of the tympanic bulla, and continued postnatal emergence of the premolars and molars from their alveoli. Disarticulated skulls suggest a degree of rostral kinesis in this genus. Phylogenetic analysis of the largest assembled supermatrix of Mysticeti (n =138 OTUs; four archaeocetes, 10 odontocetes, 124 mysticetes; 391 morphological and 27,225 molecular characters) confirms placement of Coronodon as the earliest diverging lineage of Mysticeti under equally weighted analyses whereas implied weighting places Coronodon and similar taxa outside Neoceti, prompting a review of character transformations at the base of Neoceti.
Comments: This is one of two monographs I published this year (I am very tired). This one started out in January 2022: we had an invitation to submit an article to a special volume, and we were initially going to publish the results of an analysis we reported on at the 2018 SVP meeting in Albuquerque, NM - reporting several new specimens of the toothed baleen whale Coronodon along with new phylogenetic analyses and a study of microwear in these whales in order to evaluate hypotheses of feeding behavior in early Mysticeti. Well, by July 2022 the descriptions were nearly completed and everything else had kind of ballooned and we realized we needed at least two different manuscripts - by October, the deadline, we realized we needed three different manuscripts as the descriptive paper (on its own) was already going to be a monographic-length work. As it turned out, we assumed because the journal was online and open access that there wouldn't be page limits, and the descriptive paper ended up being four times longer than the page limit - so we submitted it to PeerJ instead.
This study is the culmination of about five years of observations, but really only one year of writing. There's nothing much else research-wise that I spent doing in 2022, with the exception of the even-longer monograph published seven months later (see below!) and a cute 'little' paper coming out early 2024. This paper reports the first growth series for any toothed mysticete, names two new species of Coronodon from the Chandler Bridge Formation, Coronodon planifrons and Coronodon newtonorum. The latter was based on a manuscript name used by the late Al Sanders - 'newtoni' - except that we didn't know which member of the Newton family (who found the fossil - ChM PV 2778) - was to be honored by the name. So, at the last minute - literally during page proofs - we changed the name to 'newtonorum'.
We published this in April, and presented data from one of our two follow-up papers at this year's SVP meeting in Cincinatti, which was well-received - and as soon as the holidays are over, the three of us are going to get it wrapped up and start finalizing the tooth wear manuscript as well.

The early diverging, dolphin-sized, cetacean clade Xenorophidae are a short-lived radiation of toothed whales (Odontoceti) that independently evolved two features long thought to be odontocete synapomorphies: the craniofacial and cochlear morphology underlying echolocation and retrograde cranial telescoping (i.e., posterior migration of the viscerocranium). This family was based on Xenorophus sloanii, which, for the past century, has been known only by a partial skull lacking a braincase and tympanoperiotics, collected around 1900 from the Ashley Formation (28–29 Ma, Rupelian) near Ladson, South Carolina. A large collection of new skulls and skeletons (ChM PV 5022, 7677; CCNHM 104, 168, 1077, 5995) from the Ashley Formation considerably expands the hypodigm for this species, now the best known of any stem odontocete and permitting evaluation of intraspecific variation and ontogenetic changes. This collection reveals that the holotype (USNM 11049) is a juvenile. Xenorophus sloanii is a relatively large odontocete (70–74 cm CBL; BZW = 29–31 cm; estimated body length 2.6–3 m) with a moderately long rostrum (RPI = 2.5), marked heterodonty, limited polydonty (13–14 teeth), prominent sagittal crest and intertemporal constriction, and drastically larger brain size than basilosaurid archaeocetes (EQ = 2.9). Dental morphology, thickened cementum, a dorsoventrally robust rostrum, and thick rugose enamel suggest raptorial feeding; oral pathology indicates traumatic tooth loss associated with mechanically risky predation attempts. Ontogenetic changes include increased palatal vomer exposure; fusion of the nasofrontal, occipito-parietal, and median frontal sutures; anterior lengthening of the nasals; elaboration of the nuchal crests; and blunting and thickening of the antorbital process. The consistent deviation of the rostrum 2–5° to the left and asymmetry of the palate, dentition, neurocranium, mandibles, and vertebrae in multiple specimens of Xenorophus sloanii suggest novel adaptations for directional hearing driven by the asymmetrically oriented pan bones of the mandibles. A second collection consisting of a skeleton and several skulls from the overlying Chandler Bridge Formation (24–23 Ma, Chattian) represents a new species, Xenorophus simplicidens n. sp., differing from Xenorophus sloanii in possessing shorter nasals, anteroposteriorly shorter supraorbital processes of the frontal, and teeth with fewer accessory cusps and less rugose enamel. Phylogenetic analysis supports monophyly of Xenorophus, with specimens of Xenorophus simplicidens nested within paraphyletic X. sloanii; in concert with stratigraphic data, these results support the interpretation of these species as part of an anagenetic lineage. New clade names are provided for the sister taxon to Xenorophidae (Ambyloccipita), and the odontocete clade excluding Xenorophidae, Ashleycetus, Mirocetus, and Simocetidae (Stegoceti). Analyses of tooth size, body size, temporal fossa length, orbit morphology, and the rostral proportion index, prompted by well-preserved remains of Xenorophus, provide insight into the early evolution of Odontoceti.
Comments: I have not poured this much effort into any other project. I worked on this beast for five years, with much of the work being done in 2020.

Burin et al.: The dynamic adaptive landscape of cetacean body size.
Adaptive landscapes are central to evolutionary theory, forming a conceptual bridge between micro- and macroevolution. Evolution by natural selection across an adaptive landscape should drive lineages toward fitness peaks, shaping the distribution of phenotypic variation within and among clades over evolutionary timescales. The location and breadth of these peaks in phenotypic space can also evolve, but whether phylogenetic comparative methods can detect such patterns has largely remained unexplored. Here, we characterize the global and local adaptive landscape for total body length in cetaceans (whales, dolphins, and relatives), a trait that spans an order of magnitude, across their -million-year evolutionary history. Using phylogenetic comparative methods, we analyze shifts in long-term mean body length and directional changes in average trait values for 345 living and fossil cetacean taxa. Remarkably, we find that the global macroevolutionary adaptive landscape of cetacean body length is relatively flat, with very few peak shifts occurring after cetaceans entered the oceans. Local peaks are more numerous and manifest as trends along branches linked to specific adaptations. These results contrast with previous studies using only extant taxa, highlighting the vital role of fossil data for understanding macroevolution. Our results indicate that adaptive peaks are dynamic and are associated with subzones of local adaptations, creating moving targets for species adaptation. In addition, we identify limits in our ability to detect some evolutionary patterns and processes and suggest that multiple approaches are required to characterize complex hierarchical patterns of adaptation in deep time.

Carrasco et al.: Ecological traits of the franciscana dolphin (Pontoporia blainvillei) from the Late Pleistocene to the present days based on stable isotope analysis.
Late Pleistocene fossils from southern Brazil provide a unique opportunity to study the diet and habitat use of the extant franciscana dolphin (Pontoporia blainvillei) prior to human influence. Here, we subject 19 fossil and 21 contemporary specimens to stable carbon and oxygen isotope analysis (after first ruling out diagenesis via Fourier transform infrared spectroscopy). The two groups differ in their isotopic composition (PERMANOVA, p < .01), with fossils yielding more disparate δ13C values and recent samples more disparate δ18O values. Their isotopic niches show an overlap of ~20%, with that of the fossils being slightly wider (SEAc = 2.25‰2 versus 1.84‰2). We attribute these differences to impoverished modern fish communities and temporal changes in freshwater influx.

Collareta et al.: Polyplacophoran Feeding Traces on Mediterranean Pliocene Sirenian Bones: Insights on the Role of Grazing Bioeroders in Shallow-Marine Vertebrate Falls.
Chitons (Polyplacophora) include some of the most conspicuous bioeroders of the present-day shallow seas. Abundant palaeontological evidence for the feeding activity of ancient chitons is preserved in the form of radular traces that are usually found on invertebrate shells and hardgrounds. We report on widespread grazing traces occurring on partial skeletons of the extinct sirenian Metaxytherium subapenninum from the Lower Pliocene (Zanclean) of Arcille (Grosseto Province, Tuscany, Italy). These distinctive ichnofossils are described under the ichnotaxonomic name Osteocallis leonardii isp. nov. and interpreted as reflecting substrate scraping by polyplacophorans. A scrutiny of palaeontological literature reveals that similar traces occur on fossil vertebrates as old as the Upper Cretaceous, suggesting that bone has served as a substrate for chiton feeding for more than 66 million years. Whether these bone modifications reflect algal grazing, carrion scavenging or bone consumption remains unsure, but the first hypothesis appears to be the most parsimonious, as well as the most likely in light of the available actualistic data. As the role of bioerosion in controlling fossilization can hardly be overestimated, further research investigating how grazing organisms contribute to the biostratinomic processes affecting bone promises to disclose new information on how some marine vertebrates manage to become fossils.

Coombs et al.: Drivers of morphological evolution in the toothed whale jaw.
Toothed whales (odontocetes) emit high-frequency underwater sounds (echolocate)—an extreme and unique innovation allowing them to sense their prey and environment. Their highly specialized mandible (lower jaw) allows high-frequency sounds to be transmitted back to the inner ear. Echolocation is evident in the earliest toothed whales, but little research has focused on the evolution of mandibular form regarding this unique adaptation. Here, we use a high-density, three-dimensional geometric morphometric analysis of 100 living and extinct cetacean species spanning their ∼50-million-year evolutionary history. Our analyses demonstrate that most shape variation is found in the relative length of the jaw and the mandibular symphysis. The greatest morphological diversity was obtained during two periods of rapid evolution: the initial evolution of archaeocetes (stem whales) in the early to mid-Eocene as they adapted to an aquatic lifestyle, representing one of the most extreme adaptive transitions known, and later on in the mid-Oligocene odontocetes as they became increasingly specialized for a range of diets facilitated by increasingly refined echolocation. Low disparity in the posterior mandible suggests the shape of the acoustic window, which receives sound, has remained conservative since the advent of directional hearing in the aquatic archaeocetes, even as the earliest odontocetes began to receive sounds from echolocation. Diet, echolocation, feeding method, and dentition type strongly influence mandible shape. Unlike in the toothed whale cranium, we found no significant asymmetry in the mandible. We suggest that a combination of refined echolocation and associated dietary specializations have driven morphology and disparity in the toothed whale mandible.

The earliest fossil cetaceans (archaeocetes) dramatically shifted the shape and articulation of the pelvis and hindlimbs during the land-to-sea transition. Archaeocetes were mostly semi-aquatic “walking whales” that used powerful hindlimbs to walk on land and swim to reach new aquatic sources of food. However, skeletons of the latest diverging lineages of archaeocetes, the basilosaurids, showed that the pelvis initially lost articulation with the sacrum, and hindlimbs were reduced and encased within the body wall. Consequently, basilosaurids were no longer able to bear their weight on land and probably had a different mating strategy compared to the other archaeocetes. Basilosaurid mating behaviors were probably consistent with those of modern cetaceans, including lateral- and ventral-facing copulation. Moreover, a pelvic girdle that was no longer constrained by vertebral and limb attachments likely freed fetal development from size constraints at birth, allowing for the birth of large fetuses. This study reports new data showing growth of the pelvis with age in modern bowhead whales (Balaena mysticetus) and their implications for left-right asymmetry and sex difference in pelvic dimensions among modern cetaceans. Reproductive structures present in modern cetaceans and artiodactyls were probably present in archaeocetes, including pelvic attachment of muscles associated with erection and mobility of the penis, the ischiocavernosus, in males and the clitoris of females. Within females, transverse folds along the vaginal canal are present in some terrestrial artiodactyls, modern cetaceans, and probably archaeocetes. Vaginal folds were probably exapted to assist in successful aquatic copulation in all fossil and modern cetaceans as they may protect some sperm from the lethal effects of sea water. Taken together, shifts in the pelvic girdle of cetaceans occurred over 40 million years ago and probably required changes in mating behaviors that were consistent with those seen in modern cetaceans.

Coste et al.: A new fossil dolphin with tusk-like teeth from New Zealand and an analysis of procumbent teeth in fossil cetaceans.
Studies involving anatomical description and taxonomy of fossil odontocetes offer insights into their evolutionary history and diversity. This study analyses tusk-like teeth in odontocetes including the description of a new species, Nihoroa reimaea, from the Waitaki valley, North Otago, New Zealand. Dental features of Nihoroa reimaea, a gracile, longirostrine odontocete with procumbent tusk-like anterior teeth and slightly denticulate cheek teeth, are described in detail. A comparative analysis of tusk-like teeth from New Zealand specimens and from elsewhere in the world was performed allowing a classification of tusk-like teeth in odontocetes and highlighting the differences between true tusks and rooted procumbent teeth. Correlation analyses revealed significant associations between rostrum proportions and tooth crown morphology. This study contributes to the understanding of tusk-like teeth and illuminates their significance in odontocete evolution. Nihoroa reimaea expands our knowledge of fossil cetaceans and highlights the importance of New Zealand's exceptional fossil record of odontocetes with tusk-like teeth.

All extant toothed whales (Cetacea, Odontoceti) are aquatic mammals with homodont dentitions. Fossil evidence from the late Oligocene suggests a greater diversity of tooth forms among odontocetes, including heterodont species with a variety of tooth shapes and orientations. A new fossil dolphin from the late Oligocene of New Zealand, Nihohae matakoi gen. et sp. nov., consisting of a near complete skull, earbones, dentition and some postcranial material, represents this diverse dentition. Several preserved teeth are horizontally procumbent, including all incisors and canines. These tusk-like teeth suggest adaptive advantages for horizontally procumbent teeth in basal dolphins. Phylogenetic analysis places Nihohae among the poorly constrained basal waipatiid group, many with similarly procumbent teeth. Features of N. matakoi such as its dorsoventrally flattened and long rostrum, long mandibular symphysis, unfused cervical vertebrae, lack of attritional or occlusal wear on the teeth and thin enamel cover suggest the rostrum and horizontally procumbent teeth were used to injure and stun prey though swift lateral head movements, a feeding mode that did not persist in extant odontocetes.
Comments: The incredible tusked waipatiid is finally named! This was by far and away the most spectacular of the unpublished fossils in the Fordyce collection at University of Otago, and I'm very glad that it was published this year. Now known to the world as Nihohae matakoi, this Waipatia-like dolphin is hypothesized to have used its teeth a bit like a sawfish or sawshark with lateral sweeping movements of the neck and head to impale fish on its laterally projecting tusks.

Davydenko et al.: Gross and microscopic anatomy of a tibia tentatively attributed to a cetacean from the Middle Eocene of Europe, with a note on the artiodactyl Anoplotherium and on the perissodactyl Lophiodon.
Transition of cetaceans from a semi-aquatic to a fully aquatic lifestyle during the Eocene involved numerous transformations of their anatomy, including reduction of the hind limbs. The earliest fully aquatic cetaceans retained externally protruding, possibly functional hind limbs. Being extremely rare, every find of archaeocete hind limb elements can improve our knowledge on the evolution of locomotor strategies in cetaceans. Here, we describe the anatomy and inner microstructure of an isolated tibia fragment, that has been found in the Middle Eocene of Helmstedt, present-day Germany, with comments on its affinities and function. Assignment to an archaeocete is most probable, as other, presently known taxa could be excluded, because of the shape of the distal epiphyseal side and the internal architecture. Nevertheless, it cannot be ruled out with complete certainty that this isolated tibia belongs to a taxon that is new, or is at this moment unknown from coeval sediments of Europe. Well-developed, distally extended tibial crests could indicate the capability of movements with the foot and thus feet-induced aquatic locomotion. The tibia fragment may belong to a large member of semi-aquatic cetaceans (Protocetidae) or to a member of Basilosauridae, early fully aquatic whales. It shares gross and inner morphological features with tibiae of Protocetidae. In this case, it could be from a large protocetid, certainly more than 4 m long, and would be the first protocetid find in Europe as well as the northernmost record of this family in the world. Alternatively, the tibia may belong to a representative of Basilosauridae, which have been found in different parts of Europe, including the region of Helmstedt. If so, it would imply that early basilosaurids had large, protocetid-like hind limbs. This record shows that large quadrupedal cetaceans inhabited the seas of Northwestern Europe by the late Middle Eocene. Not-related finds of our studies are that the terrestrial lifestyle of the artiodactyl Anoplotherium is confirmed by the inner structure of its long bones, and that some species of the perissodactyl Lophiodon were most probably semi-aquatic, as shown by the inner microstructure of the tibia.

There are a few mammalian lineages secondarily adapted to life in water throughout their evolutionary history. Of them, only sirenians and cetaceans evolved as fully aquatic organisms. This transition was accompanied by changes in swimming mode, from foot paddling to tail-powered propulsion, with the forelimbs acting as efficient hydrofoils and the gradual loss of external hindlimbs. Here, we describe an isolated limb from the Middle Eocene of Ukraine, 43–42 Mya, identified as a hindlimb of a fully aquatic cetacean and being the earliest cetacean reported from Europe. It is represented by flattened, jointly articulated bones, identified as the tibia and fibula with a partly reduced knee joint and loose connection to the pes, and by flattened phalanges. This anatomy reveals a hitherto only presumed cetacean morphotype, showing that some of the early fully aquatic cetaceans were four-legged animals with functional hindlimbs that could be involved in advanced styles of swimming. They used either body undulation or lift-based propulsion powered by the tail or feet and could also use a transitional swimming style combining these modes.

Davydenko et al.: Diverse bone microanatomy in cetaceans from the Eocene of Ukraine further documents early adaptations to fully aquatic lifestyle.
Basilosauridae, fully aquatic archaeocetes from the Eocene, had osteosclerotic or pachyosteosclerotic structure of ribs and, sometimes, other bones. Such a structure is far different from osteoporotic-like bones of modern cetaceans. A microanatomical and histological study was conducted on axial and limb skeleton of several basilosaurid specimens assigned to the genus Basilotritus, from Bartonian (late middle Eocene) deposits of Ukraine, remarkable for its pachyostotic bones. The postcranial skeleton of these specimens is a complex mosaic of diverse types of bone structure, which include pachyosteosclerotic, osteosclerotic and cancellous elements. The vertebrae have a pachyostotic layered cortex reaching its greatest thickness in the lumbar region. This cortex was strongly vascularized, and its layered structure is due to concentric circles mostly made by longitudinal vascular canals, in addition to cyclical growth lines. Heavy bones are concentrated in the dorsal and ventral areas. Swollen distal ends of thoracic ribs are interpreted as serving as ballast in the ventral area, as also previously proposed for Basilosaurus cetoides. Cortical bone tissue in vertebrae and ribs showed signs of intensive resorption and remodeling. This indicates the use of the axial skeleton not only for buoyancy control but also possibly for calcium and phosphorus recycling.

The form and function of the sacrum are of great relevance to understand the evolution of locomotion in tetrapods because it is a key piece of the vertebrate skeleton. The sacrum connects the caudal and presacral regions of the vertebral column and the hindlimbs through the pelvis. Here, we investigate sacrum shape evolution in pinnipeds (Carnivora: Pinnipedia) in relation to terrestrial mammalian carnivorans (fissipeds), and we include crown and stem taxa to quantify the morphological changes they experience in relation to the aquatic environment they inhabit. We use 3D geometric morphometric methods to explore the morphological variability and disparity of the sacrum in a set of terrestrial and aquatic carnivoran species. Our results show that the morphology of the sacrum of each pinniped family is remarkably different and that these differences may be related to the aquatic mode of locomotion (pectoral or pelvic oscillation), the use of hindlimbs to support body weight on land (otariids in contrast with phocids), and the presence or absence of a functional tail. In addition, disparity-through-time analyses indicate that the sacrum of pinnipeds is less constrained than that of fissipeds, which suggests a gravitational origin of such constraints in fissipeds. In conclusion, our results give further support to the important role played by this skeletal structure in the locomotory adaptations of mammals.

Prospecting fieldwork in a new fossiliferous locality of the marine Early Miocene Gaiman Formation (Patagonia, Argentina) results in the discovery of abundant fossil cetaceans (n = 29). We analyse stratigraphic, sedimentologic, palaeontologic, and taphonomic data with the aim of understanding the factors that control the preservation and distribution of these specimens. The cetacean sample is dominated by small odontocetes exhibiting five different preservation categories. Based on sedimentological features, three distinct facies were recognised in the stratigraphic section. Stratigraphically condensed deposits of facies C, related to a ravinement surface and subsequent transgressive deposits, enclose 52% of the specimens, grading from isolated postcranial elements to articulated and associated postcranial and cranial elements. In contrast, facies A and B exhibit predominantly isolated postcranial elements. Facies C shows higher degrees of corrosion, encrustation and fragmentation than those specimens from facies A and B, reflecting differences in preservation quality among the facies. The higher abundance and evidence of long exposure times recorded for facies C seems to be resulted mostly from physico-chemical factors after stratigraphic condensation. Finally, intrinsic biological features and ecological preferences of the specimens could explain the dominance of small-sized odontocetes in a restricted inner shelf to lower shoreface environmental setting.
Comments: I really need to read this one, but it was very satisfying to see conceptual figures like the one above like those that I published in my master's thesis on the taphonomy of the Purisima Formation.

Gaetan et al.: A new squaloziphiid-like odontocete from the Early Miocene of Patagonia expands the
cetacean diversity in the southwestern Atlantic Ocean.
The Early Miocene Gaiman Formation, from Argentina, houses a taxonomic variety of odontocetes, mainly dominated by platanistoids and physeteroids. In this work, we describe a new medium-sized odontocete, Crisocetus lydekkeri gen. et sp. nov., based on a partial skull, which expands the diversity of odontocetes in the Early Miocene beds of Patagonia (Argentina). The phylogenetic analyses placed C. lydekkeri within the crown Odontoceti, stemward to delphinidans, ziphiids, and physeteroids. Crisocetus lydekkeri displays a set of characters that are shared with the members of the Squaloziphiidae family (i.e., Squaloziphius emlongi and Yaquinacetus meadi), such as a massive postglenoid process of the squamosal, ventrally longer than the posttympanic process and the exoccipital; apex of postglenoid process anteroposteriorly longer than transversely thick; presence of a deep emargination by a neck muscle fossa on the posterior end of the zygomatic process; basioccipital crests forming an angle of circa 70–90°; and a basioccipital width wider than 51% of the skull width in ventral view. Thus, we recognize a squaloziphiid-like morphology shared by C. lydekkeri, S. emlongi, Y. meadi, and even Dolgopolis kinchikafiforo. Our phylogenetic analyses partially support the inclusion of C. lydekkeri within Squaloziphiidae, however this family is recovered with low support. Finally, the presence of C. lydekkeri and even D. kinchikafiforo in the Early Miocene of Patagonia expands the paleogeographic distribution of squaloziphiid-like forms to the southwest coast of the Atlantic Ocean, suggesting that these taxa had an almost antitropical distribution.
Comments: I really, really wish this specimen had a periotic!

[Abstract for entire Volume] This volume is a follow-on to a work published by Smithsonian Institution Scholarly Press in 2018 on the Miocene vertebrate fauna from Calvert Cliffs, Maryland, USA. Two chapters are included in this compendium, one on turtles (chelonians) and the other on toothed whales (odontocetes). It is anticipated that at least one more volume will be needed to complete the taxonomic review. Robert E. Weems details the occurrence of 19 kinds of chelonians that have been discovered in the Miocene and Pliocene marine strata of Delaware, Maryland, and Virginia, USA, 13 of them in the Calvert Cliffs. The most commonly found remains are those of an extinct sea turtle, Trachyaspis lardyi. Remains of four other marine turtles, Procolpochelys grandaeva, Lepidochelys sp., a generically indeterminate cheloniid, and a leatherback turtle (Psephophorus polygonus), are far less common. The other 14 chelonian taxa are nonmarine forms that inhabited the land, rivers, and marshes west of the Mid-Atlantic Seaboard during the Miocene. They were washed into the coastal marine environments that were then accumulating the sediments exposed today as the strata in the Calvert Cliffs. Stephen J. Godfrey and Olivier Lambert review the taxonomically diverse odontocete fauna of 29 named species. Nine of these Miocene taxa represent newly named species. Fragmentary remains hint at even greater diversity. Reviewed taxa are restricted to those known from along the Calvert Cliffs and other Miocene age deposits on the Atlantic Coastal Plain in Maryland and Virginia, USA. They range in age from approximately 22 to 8 Ma and derive from the Calvert, Choptank, and St. Marys Formations. This fauna preserves one of the most abundant and diverse assemblages of extinct toothed whales known. None of the named odontocete species included in this review are known from beyond the North Atlantic Ocean. In terms of their chronostratigraphic distribution, collectively, they range in age from the Aquitanian through the Tortonian, with the large majority occurring within the Burdigalian, Langhian, and Serravallian stages (the latter two being the most speciose). The greatest taxonomic diversity occurred during the Middle Miocene Climate Optimum, a time (ca. 17–15 Ma) when global average temperatures were as much as 4°C to 5°C above today’s average temperatures, at least for much of that interval.
Comments: Another monumental paper for 2023 on fossil odontocetes! It's been a very good year for odontocetes, with Xenorophus, simocetids, and waipatiids very well represented. This study names a bunch of new taxa, including the new shark-toothed dolphin Squalodon murdochi, the new platanistids Grimadelphis spectorum and Pomatodelphis santamaria, the new kentriodontids Herbeinodelphis nancei, Cammackacetus hazenorum, Pictodelphis kidwellae, Westmorelandelphis tacheroni, the Eoplatanista-like Caolodelphis milleri, and the utterly bizarre Enigmatocetus posidoni. The study also includes a formal revision of Calvert eurhinodelphinids, now identified as Xiphiacetus bossi, Xiphiacetus cristatus, Schizodelphis barnesi, and Schizodelphis morckhoviensis. The kentriodontid dolphin Delphinodon dividum has been redescribed and reassigned to the new genus Brevirostrodelphis, and recombined as Brevirostrodelphis dividum. In a similar vein, Liolithax pappus has been reassigned to the new genus Miminiacetus - and great new specimens are referred to this species. The kentriodontids Hadrodelphis and Macrokentriodon have been re-figured, which is excellent because the original figures are missing some views and the quality of the photographs is not great. Also, new specimens of certain odontocetes include quite a bit more material - including the first periotic for the platanistoid Araeodelphis natator. The periotic of the kentriodontid Lophocetus calvertensis has also been finally figured. Altogether, a phenomenal contribution - this study did stop short of reporting isolated periotics from Calvert Cliffs - which is assuredly a gargantuan task.

Govender and Marx: New cetacean fossils from the late Cenozoic of South Africa.
Marine sediments from the western coast of southern Africa record the origin of the Benguela Upwelling System, one of the most productive in the world. High productivity, in turn, is reflected in a diverse marine mammal fossil assemblage, comprising whales, dolphins and a phocid seal. Here, we describe new records of baleen whale (mysticete) fossils from the early Pliocene localities of Saldanha Steel, Milnerton and Langebaanweg, as well as several potentially younger specimens trawled from offshore sediments. The presence of the extinct rorquals Diunatans and Fragilicetus suggests biogeographical links with the eastern North Atlantic and, thus, potentially antitropical population structuring. The trawled specimens also include rorquals (e.g., the blue whale, Balaenoptera cf. musculus), as well as a right whale (Eubalaena) and a pygmy right whale (Caperea). The latter is the first fossil of this family every discovered in Africa, and only the seventh specimen to be reported worldwide.
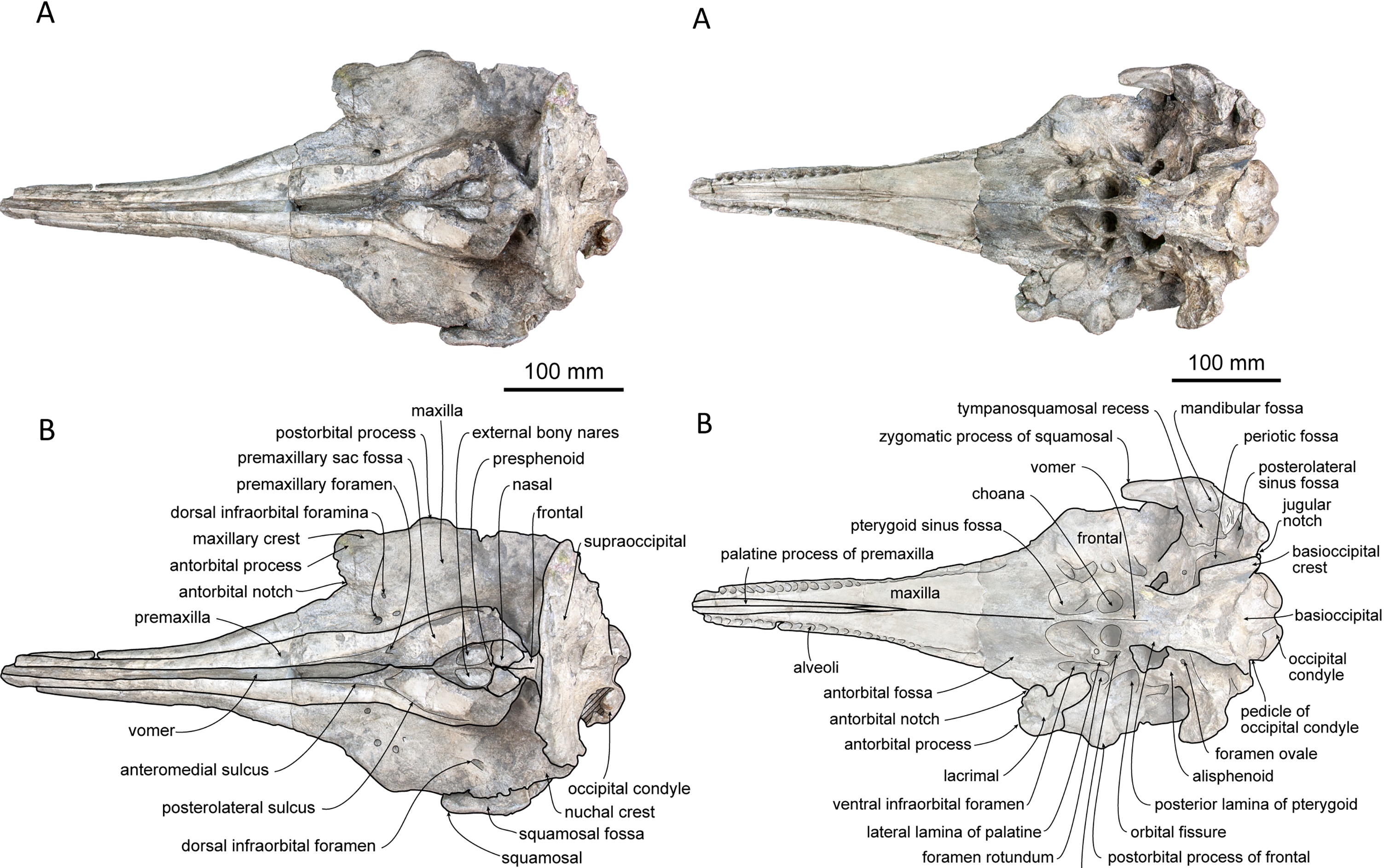
Guo and Kohno: An Early Miocene kentriodontoid (Cetacea: Odontoceti) from the western North Pacific, and its implications for their phylogeny and paleobiogeography.
So–called ‘kentriodontids’ are extinct dolphin–like odontocetes known from the Early to Late Miocene worldwide. Although recent studies have proposed that they were monophyletic, their taxonomic relationships still remain controversial. Such a controversy exists partly because of the predominance of primitive morphologies in this taxon, but the fact is that quite a few ‘kentriodontids’ are known only from fragmentary skulls and/or isolated periotics. A new ‘kentriodontid’ Platysvercus ugonis gen. et sp. nov. is described based on a nearly complete skull from the upper Lower Miocene Sugota Formation, Akita Prefecture, northern Japan. Based on the phylogenetic analysis of P. ugonis described here, the monophyly of the ‘kentriodontids’ is confirmed, and it is recognized as the superfamily Kentriodontoidea. This new superfamily is subdivided into two families as new ranks: Kentriodontidae and Lophocetidae. Based on the paleobiogeographic analysis of the Kentriodontoidea, their common ancestor emerged in the North Pacific Ocean and spread over the Northern Hemisphere. Initial diversification of the Kentriodontidae in the North Pacific Ocean and the Lophocetidae in the North Atlantic Ocean was recognized as a vicariance event. The diversification and extinction of the Kentriodontoidea could have been synchronously influenced by climate events during the Middle Miocene.

Hautier et al.: From teeth to pad: tooth loss and development of keratinous structures in sirenians.
Sirenians are a well-known example of morphological adaptation to a shallow-water grazing diet characterized by a modified feeding apparatus and orofacial morphology. Such adaptations were accompanied by an anterior tooth reduction associated with the development of keratinized pads, the evolution of which remains elusive. Among sirenians, the recently extinct Steller's sea cow represents a special case for being completely toothless. Here, we used μ-CT scans of sirenian crania to understand how motor-sensor systems associated with tooth innervation responded to innovations such as keratinized pads and continuous dental replacement. In addition, we surveyed nine genes associated with dental reduction for signatures of loss of function. Our results reveal how patterns of innervation changed with modifications of the dental formula, especially continuous replacement in manatees. Both our morphological and genomic data show that dental development was not completely lost in the edentulous Steller's sea cows. By tracing the phylogenetic history of tooth innervation, we illustrate the role of development in promoting the innervation of keratinized pads, similar to the secondary use of dental canals for innervating neomorphic keratinized structures in other tetrapod groups.

Helmy et al.: Valley of Whales, Fayum oasis, Egypt: an Eocene window in the evolution of Cetaceans.
The present-day Mediterranean Sea is the remnant of the Tethys Ocean a former tropical body of saltwater that separated the continental blocks of Laurasia from Gondwana during much of the Mesozoic and early Cenozoic Era. The Tethys Sea extended far south into Gondwana (i.e., northern Africa) and gradually retreated north depositing thick layers of fossiliferous marine sediments (sandstone, limestone and shale) of early Palaeogene age in the area of northern Egypt. This slow regression through millions of years created a northward moving shoreline separating between terrestrial and marine environments. Palaeomagnetic studies (Lotfy and Van der Voo 2007) suggest that this retreating shoreline crossed equatorial palaeo-latitudes in a tropical setting. A plethora of marine invertebrate and vertebrate faunal remains (including numerous skeletons of early whales, Fig. 1) as well as mangrove roots, were deposited in the shallow marine near-shore sediments at the Valley of Whales (Wadi Al-Hitan) area in the Western Desert of Egypt.

Taxonomic identification of whale bones found during archaeological excavations is problematic due to their typically fragmented state. This difficulty limits understanding of both the past spatio-temporal distributions of whale populations and of possible early whaling activities. To overcome this challenge, we performed zooarchaeology by mass spectrometry on an unprecedented 719 archaeological and palaeontological specimens of probable whale bone from Atlantic European contexts, predominantly dating from ca 3500 BCE to the eighteenth century CE. The results show high numbers of Balaenidae (many probably North Atlantic right whale (Eubalaena glacialis)) and grey whale (Eschrichtius robustus) specimens, two taxa no longer present in the eastern North Atlantic. This discovery matches expectations regarding the past utilization of North Atlantic right whales, but was unanticipated for grey whales, which have hitherto rarely been identified in the European zooarchaeological record. Many of these specimens derive from contexts associated with mediaeval cultures frequently linked to whaling: the Basques, northern Spaniards, Normans, Flemish, Frisians, Anglo-Saxons and Scandinavians. This association raises the likelihood that early whaling impacted these taxa, contributing to their extirpation and extinction. Much lower numbers of other large cetacean taxa were identified, suggesting that what are now the most depleted whales were once those most frequently used.

Kampouridis et al.: The Eocene–Oligocene Vertebrate Assemblages of the Fayum Depression, Egypt.
The Fayum has yielded one of the oldest and richest records of fossil mammals from Africa. Today, the Fayum Depression represents an oasis in the Western Desert of Egypt, south of Cairo, and contains several localities that are world renowned for their diverse Eocene to Oligocene vertebrate assemblages. The fossil sites of the Fayum area have provided numerous extraordinarily well-preserved vertebrate remains including complete skulls and partial skeletons of turtles, crocodiles, birds, and mammals. Thus, the Fayum Depression has shed light onto the evolution and biogeography of vertebrates during the Paleogene leading to the establishment of two new orders, several new families and subfamilies, and numerous new species of mammals. In recent years, these fossils have provided clues about ecological aspects of some groups using new methods such as stable isotope analysis and µCT scanning. Despite the fact that the Fayum Depression represents a historical excavation site that has been excavated and studied by numerous famous palaeontologists since the nineteenth century, the area continues to provide new insights into the evolution of mammals during the Eocene to Oligocene, greatly improving our understanding of early Cenozoic vertebrate evolution. This chapter presents a historical review of the excavations in the Fayum Depression and a taxonomic overview of its fossil fauna. Special focus was placed on the taxonomy and, where applicable, ecology of terrestrial mammals from the historical Paleogene Fayum localities.

Kimura and Hasegawa: Additional rostrum fragments of the holotype of Miophyseter chitaensis.
Miophyseter chitaensis is a primitive physeteroid, which were found from the lower Miocene Morozaki
Group, central Japan. Kimura and Hasegawa(2022)described the cranium of the holotype (TMNH10785) which lacks most of the rostrum. In this short note, we describe three additional fragments of the rostrum of TMNH10785. The rostrum fragments were found from the matrix of the holotype of M. chitaensis. One of the fragments perfectly fits the already described rostrum fragment of the holotype, clearly suggesting that the fragments originated from the same individual of TMNH10785. An additional part of the rostrum fragments clearly suggests that M. chitaensis had a bottleneck-shaped rostrum.

This study describes a previously undescribed fossil physeterid from the Pleistocene of Tokyo. This fossil physeterid was recovered from the Early Pleistocene, Fukushima Formation (ca. 1.60‒1.77 Ma), Kazusa Group in 1971 and has long been known as “Hino Kujira” without detailed descriptions. The specimen consists of fragments of the left maxilla, and we estimate the original size of the maxilla can reach at least 3 m long, suggesting an individual at least 11.5 m long. The maxilla is dorsoventrally thick (eroded, but at least 188 mm), and such a thick maxilla clearly differs from that of mysticetes but is similar to that of the modern sperm whale, Physeter macrocephalus. In addition, the average size of each pore on the spongy bone under the compact bone layer is prominent (10 to 15 mm in diameter), consistent with the macroporous structure of the maxilla of modern Physeter macrocephalus. We then provisionally make the taxonomic assignment of this Pleistocene specimen as Physeter sp. Besides, we review seventy-three documented fossil cetaceans from the Pleistocene of Japan, suggesting that the known paleodiversity of the Japanese Pleistocene cetaceans (12 genera) is much lower than that of modern composition (26 genera). This evident discrepancy likely results from the research effort that currently only produces an inadequately known fossil record of cetaceans.

Carbonate concretions occur in sedimentary rocks of widely varying geological ages throughout the world. Recently, more than 100 gigantic carbonate concretions with diameters ranging from 1 to 9 m have been identified along the Unosaki coast of Oga Peninsula, Akita Prefecture, Japan. The formation process of such gigantic concretions, some of which along the Unosaki coast contain whale bones, remains uncertain. A mineral composition analysis reveals that the major mineral of the concretions is dolomite. Considering the location of dolomite precipitation, their composition implies that the concretions were formed in a reducing environment in which sulfate ions were removed. Stable carbon and oxygen isotopic analysis reveals that the CaCO3 of whale bone and concretions contains light δ13C and heavy δ18O, suggesting that whale organic matter contributed to the formation of the concretions. The gigantic carbonate concretions were presumably formed by the accumulation and burial of whale carcasses with high sedimentation rates, and subsequent reaction of carbon decomposed by benthic and microbial activity with seawater.
Remarks: I can't read the majority of this article, but I will note that there are large dolomitic whale-bearing concretions in the Purisima Formation at Point Reyes (Marin County) - sometimes these concretions are up to about 5 meters across. Dick Hilton described to me a visit he took to the beach there (sometime in the 1960s I believe) after a large storm had stripped off much of the sand, and there was a 5-6 meter wide donut-shaped concretion with a baleen whale skeleton, that had been preserved in a C-shape.

Extant beaked whales (family Ziphiidae) are deep diving suction feeders and none of them can be considered as a permanent resident of the shallow southern North Sea. The rich fossil record of ziphiids from Neogene deposits of this area is thus surprising. However, chronostratigraphic intervals of most recorded taxa remain poorly constrained, preventing from assessing the evolution of their local diversity. In this work, we describe a new ziphiid cranium from the Neogene of Antwerp (north of Belgium), which is attributed to Caviziphius aff. C. altirostris. Sediment samples were extracted from 15 fossil ziphiid cranial remains from this area (including the one described herein), referred to eight species. The samples were analysed for their palynological content, leading to improved chronostratigraphic ranges for several species. Seven to eight ziphiid species from the southern North Sea, all from the Messapicetus clade, are proposed to originate from a Serravallian to Tortonian (late Middle to early Late Miocene) interval, and three to six more precisely from the mid- to late Tortonian. Added to the fossil record of other regions, these results point to a Late Miocene radiation of members of the Messapicetus clade, possibly related to the synchronous worldwide decline of several hyper-longirostrine dolphin clades.

Lambert et al.: A new platyrostrine sperm whale from the Early Miocene of the southeastern Pacific (East Pisco Basin, Peru) supports affinities with the southwestern Atlantic cetacean fauna.
Contrasting with their suction feeding modern relatives in the families Kogiidae and Physeteridae, Miocene physeteroids display a broad range of feeding strategies. Despite the continuous improvements of the fossil record, the transition from the earliest sperm whales to suction feeding forms as well as the once prominent macroraptorial forms remains poorly understood. In the present work, we investigate a partial sperm whale skull from Lower Miocene (Burdigalian) strata of the Chilcatay Formation of the East Pisco Basin, along the southern coast of Peru. Based on this specimen, we describe a new species in the genus Diaphorocetus Ameghino, 1894, which was previously known only by the holotype of Diaphorocetus poucheti (Moreno, 1892) from a roughly synchronous unit in Patagonia (Argentina). Differing from the latter in its smaller cranial dimensions, higher tooth count, and minor differences in the position of facial foramina, the new species Diaphorocetus ortegai n. sp. confirms a key character of D. poucheti, the marked dorsoventral flattening of the maxillary portion of the rostrum. Such cranial proportions suggest that, compared to other physeteroids, D. poucheti and D. ortegai n. sp. were more efficient at performing fast lateral sweeps of their rostra to capture small- to medium-sized prey items with their proportionally small teeth. Recovered as stem physeteroids in our phylogenetic analysis, these sister species contribute to the ecomorphological disparity of sperm whales during the Early Miocene, but without displaying any of the cranial and dental changes occurring in later, macroraptorial and suction feeding sperm whales. The description of a new species of Diaphorocetus from southern Peru increases the similarities between the toothed whale faunas from the local Chilcatay Formation and the Gaiman and Monte Leon formations of Argentinian Patagonia, pointing not only to dispersal routes between the southeastern Pacific and southwestern Atlantic during the Burdigalian, but also to relatively similar ecological settings along the coasts of Peru and Patagonia at that time.

Lambert et al.: First eurhinodelphinid dolphin from the Paratethys reveals a new family of specialised echolocators.
Eurhinodelphinids are a family of extremely long-snouted dolphins that once was widespread across the North Atlantic realm, but so far has not been recorded from the epicontinental Paratethys Sea. Here, we describe a new specimen of Xiphiacetus cristatus, including the cranium, left ear bones and a partial mandible, from the Middle Miocene (Badenian) of Austria. Our new fossil is the first record of this species outside the North Atlantic proper, and the first unequivocal record of eurhinodelphinids from the Paratethys. Its presence suggests a local invasion via the Mediterranean, and corroborates the persistence of an active marine gateway between the Mediterranean and the Central Paratethys throughout the Middle Miocene. Measurements of the bony labyrinth reveal that X. cristatus likely employed narrow-band high-frequency echolocation, making eurhinodelphinids only the second extinct odontocete family with this trait. Marked non-orthogonality of the semicircular canals suggests that X. cristatus was unsuited to rapid head movements, and thus more likely a benthic forager than a snap feeder.

Leon et al.: New fossil Cetaceans from the Middle Miocene of Cuba.
Remains of cetaceans, particularly odontocetes, are scarce in the Cuban fossil record. Here we describe the second odontocete (toothed whale) record from the Neogene of Cuba and extends the temporal distribution of this group in the Insular Caribbean into the Middle Miocene. This account is based on two isolated teeth found in limestones of the Güines Formation that crop out in a quarry near the town of Guanábana, Matanzas City, northwestern Cuba. The specimens are referred to the superfamily Physeteroidea incertae sedis. These findings, along with other fossils recovered from other Early and Middle Miocene localities in the region support the hypothesis of a complex, diverse marine vertebrate fauna on the formerly called “Habana-Matanzas channel” and circum-Cuban waters during the Neogene.

Pinnipeds (seals and related species) use their whiskers to explore their environment and locate their prey. Today they live mostly in marine habitats and are adapted for a highly specialised amphibious lifestyle with their flippers for locomotion and a hydrodynamically streamlined body. The earliest pinnipeds, however, lived on land and in freshwater habitats, much like mustelids today. Here we reconstruct the underwater foraging behaviour of one of these earliest pinnipeds (Potamotherium), focusing in particular on how it used its whiskers (vibrissae). For this purpose, we analyse the coronal gyrus of the brain of 7 fossil and 31 extant carnivorans. This region receives somatosensory input from the head. Our results show that the reliance on whiskers in modern pinnipeds is an ancestral feature that favoured survival of stem pinnipeds in marine habitats. This study provides insights into an impressive ecological transition in carnivoran evolution: from terrestrial to amphibious marine species. Adaptations for underwater foraging were crucial for this transition.

A c. 31 m thick section straddling the fossil find of an Early Pliocene baleen whale (“Brunella”, hereafter), made in 2007 in the sedimentary fill of the Middle Ombrone Basin of Tuscany, is investigated for depositional age and environment combining palaeomagnetic, micropalaeontological (Foraminifera and calcareous nannofossils) and sedimentary facies analyses. Resting unconformably onto Late Miocene continental deposits, the Early Pliocene marine deposits include, from bottom to top, a coarse-grained wave-winnowing lag, the few metres-thick fossiliferous sandstone bedset from which Brunella was unearthed, and several metres of clays. The stratigraphic organisation of these deposits indicate deposition in a deepening upward inner shelf environment. Successful isolation of characteristic remanent magnetisation and calcareous nannofossil content indicate the investigated marine section was deposited during the interval of polarity Chron C3n.2n corresponding to the basal part of the Mediterranean nannofossil zone MNN13 (between Helicosphaera sellii Base common and the Amaurolithus primus Top) and allow estimating the depositional age of Brunella to c. 4.6 Ma. Sedimentary facies, benthic Foraminifera association and anisotropy of magnetic susceptibility characterising the deposits that embedded Brunella suggest deposition above the fair-weather base level.

Marx et al.: Suction causes novel tooth wear in marine mammals, with implications for feeding evolution in baleen whales.
Teeth are the primary tool used by most mammals to capture and process food. Over the lifetime of an individual, they progressively wear through contact with each other (attrition) and with food (abrasion), creating distinctive patterns that reflect function and diet. Unlike their terrestrial cousins, many marine mammals capture prey via suction, which so far has not been associated with a specific wear pattern. Here, we describe two new types of tooth wear across 18 species of modern marine mammal (beaked whales, belugas, killer whales, globicephalines, and various seals) that likely stem from this behaviour: “glossowear”, which primarily affects the lingual side of the crown and plausibly records piston-like tongue movements during suction feeding; and “hydrowear”, which wraps around the sides of the crown and occurs as water is expelled from the mouth. Both wear types differ from attrition and biting-related abrasion in their surface characteristics and location on the crown. Horizontal scratches suggest a physical wear process, rather than dental erosion (acid corrosion) and tooth abfraction (microfracture). Since suction specifically exploits the liquid properties of water, physical evidence of this behaviour may help to elucidate marine mammal feeding ecology and evolution. For example, glossowear is found in the toothed ancestors of baleen whales (mammalodontids, at least one aetiocetid, and likely Mystacodon), where it suggests an important role for suction in the emergence of filter feeding. By contrast, it is absent in most long-snouted toothed whales and dolphins, indicating that these animals mostly bite, rather than suck in, their prey.

Aotearoa New Zealand today is home to nine extant species of pinniped (seals and sea lions) and 45 cetaceans (whales and dolphins), many of which are globally rare (Table 25.1; Figs 25.1, 25.2). This is slightly more than the eight pinnipeds and 43 cetaceans listed in King et al. (2009), with new discoveries like Ramari’s beaked whale, Mesoplodon eueu Carroll et al., 2021, continuing to add to this diverse local fauna. New Zealand’s marine mammal fossil assemblage is equally rich and has increased by twelve species since King et al. (2009), with a further four having been reassigned to new genera. There is enormous potential for further fossil discoveries, with research so far having focused largely on the upper Oligocene (28–23 million years ago) of the southeastern South Island. The lists of New Zealand extant and fossil marine mammals reported by King et al. (2009) are reviewed here, and updated checklists are provided.

Desmostylus is an extinct marine mammal genus that belongs to Desmostylia, a clade of extinct herbivorous mammals. While desmostylian remains are widely reported from Paleogene and Neogene marine strata of the North Pacific Rim, occurrences of the genus Desmostylus are almost entirely limited to middle Miocene strata, with only a few early Miocene records from Japan. Here we report a Desmostylus tooth from the earliest Miocene (Aquitanian) Skooner Gulch Formation in northern California, USA. This specimen exhibits cuspules around the crown, a primitive trait of the subfamily Desmostylidae, as seen in more basal branching desmostylid taxa such as Cornwallius and Ounalashkastylus, but with a high tooth crown and thickened enamel. The specimen is also diagnostically different from all other desmostylid genera, such as Cornwallius, and Ounalashklastylus. The Aquitanian age of the Skooner Gulch Formation implies that the distinctive tooth morphology of Desmostylus has persisted, largely unchanged, for more than 15 million years and that desmostylids possibly originated in western North America.

McMillan: Albert Koch's Hydrarchos: a hoax or a bona fide collection of bones.
This is the second essay of a two-part series on the life and collecting activities of Albert Koch. After Koch traveled to England where he sold his Missourium to the British Museum, the American mastodon that now stands in the Natural History Museum of London, he then went to his homeland in Germany. Koch left his family in Dresden, when he again departed for the United States to pursue some additional paleontological adventures. Following several weeks of travel, he arrived in Alabama where he excavated the remains of a large, archaeocete whale, that he named the Hydrarchos. Koch displayed the skeleton in New York, and several other eastern cities before taking it to Europe. When in Berlin, Koch was able to sell the skeleton to King Friedrich Wilhelm IV of Prussia who placed it in the Royal Museum. Soon thereafter, Koch returned to the United States and Alabama to discover a second whale skeleton. He took this skeleton to Europe where it was exhibited in several cities but, having received no offers, Koch returned with his second Hydrarchos to the United States where it was initially displayed in New Orleans, then St. Louis, and eventually Chicago. In his later years, Koch turned his attention to the Academy of Science of St. Louis where he became an active member and curator, as well as a prospector for minerals. This essay examines the final chapters of Koch’s life and his entrepreneurial showmanship tendencies versus contributions he may have made to science. This narrative is a sequel to an article published in Volume 41 Number 2 of Earth Sciences History that focused on Albert Koch’s Missourium. Together, the two essays capture the life and career of Albert C. Koch.

Merella et al.: Structured-Light 3D Scanning as a Tool for Creating a Digital Collection of Modern and Fossil Cetacean Skeletons (Natural History Museum, University of Pisa).
The Natural History Museum of the University of Pisa hosts one of the most important osteological collections of cetaceans all over Europe, as well as a conspicuous paleontological collection, including several holotypes of Archaeoceti (archaic whales), Mysticeti (baleen whales), and Odontoceti (toothed whales). In order to valorize these collections, we used 3D technologies to digitize the most relevant specimens, create an online archive, and make the resulting models accessible and shareable with the broadest audience possible through social media profiles and internet browsers. Tens of specimens were surface-scanned using a structured-light scanner, and the resulting 3D models were processed for post-production through the 3D software Blender whenever necessary. All the 3D scans were then gathered in the online repository Sketchfab, which was chosen for its user-friendly interface and common usage among museum institutions. The result is a web page that hosts 35 surface scans of extant and extinct cetacean specimens. This Sketchfab account was linked to the social media (Facebook and Instagram) profiles of the MSNUP to increase the visibility of the museum and promote the dissemination of its outstanding collections of modern and fossil cetaceans. The preliminary results of such an effort are encouraging in terms of views and online interactions. Hopefully, this effort of digitization and online archiving will soon extend to other vertebrate collections.

Quarries are areas of pivotal importance for investigation, popularization, and educational purposes in the field of geosciences. Here, we focus on three quarries of Tuscany (La Serra, Arcille, and Certaldo) that have been home to significant finds of marine vertebrate fossils. The three selected localities are representative of the many active (La Serra), inactive (Arcille), and abandoned (Certaldo) Tuscan quarries where vertebrate-bearing Pliocene deposits are exposed. Given the richness and scientific value of their paleontological content (including holotypes as well as otherwise unique specimens), the abundance of exposed sedimentary structures, and the high potential for open-air musealization, the La Serra, Arcille, and Certaldo quarries should be regarded as geosites. These quarries are thoroughly described, and several suggestions for their preservation and valorization are proposed, focusing on their importance for geotourism, with the goal of reaching out to the broadest audience possible. Many innovative dissemination tools, including 3D technologies, are now available for pursuing such an aim.

Fossils in the order Sirenia (family Dugongidae) from Santa Rosa Island, part of Channel Islands National Park in southern California, provide rare temporal and spatial links between earlier and later evolutionary forms of dugongids, and add information about their dispersal into the northeastern Pacific region. Marine sedimentary rocks containing these fossils have characteristics of both the late Oligocene to middle Miocene Vaqueros Sandstone and the early to middle Miocene Rincon formation observed elsewhere. To determine a more precise age of the fossils, marine invertebrate shells were collected from the same exposures as the sirenian fossils for chronostratigraphic assessment using strontium isotope compositions and the well-calibrated seawater strontium evolution curve. Shells used for analysis were from bivalve mollusks (Pycnodonte sp. [oyster] and Lyropecten sp. [scallop]) and crustaceans (Balanus sp. [barnacle]). Results show a wide range of 87Sr/86Sr values, indicating that shell materials experienced varying degrees of diagenetic alteration. Strontium concentrations and 87Sr/86Sr values in subsamples of Pycnodonte shell show correlations between original shell material and a secondary component having lower strontium concentrations and less radiogenic (lower) 87Sr/86Sr. In contrast, all Lyropecten shell analyses yielded a uniform 87Sr/86Sr value (0.708440±0.000010 [2× standard deviation]) over a wide range of strontium concentrations (around 900 to 1,800 micrograms per gram [µg/g]). Results for Balanus shell subsamples show a range of strontium compositional behavior between the other two types of shell. Acetic acid leachates of sandy matrix confirm that diagenetic fluids had low 87Sr/86Sr values consistent with the least radiogenic values in Pycnodonte subsamples. A simple mixing model between two calcite end-members can explain observed Pycnodonte data, although actual diagenetic processes likely involved secondary dissolution/reprecipitation or strontium ion exchange between shell material and pore fluid. Data indicate that only Lyropecten subsamples have retained their original 87Sr/86Sr compositions, resulting in a best-estimate age of 20.08±0.11 million years ago (Ma) (±95-percent confidence interval [CI]). Although Dugongidae fossils have been found in Miocene and younger sediments along the west coast of North America, the Santa Rosa Island specimens represent some of the earliest and most accurately dated sirenian fossils in the region. Chronostratigraphic results also constrain the timing of the transgressional processes represented by shallow-water (Vaqueros Sandstone) to deep-water (Rincon formation) depositional environments.

Ponomarev et al.: Pleistocene Walrus Odobenus rosmarus (L., 1758) Discovered in Northern European Russia (Pechora River)
A fragment of a walrus skull of Neopleistocene age was found at a distance of about 340 km from the mouth of the Pechora River. The skull presumably belonged to a mature male Atlantic walrus 13–14 years old. The radiocarbon date of the walrus bone shows an age outside the method’s range (>45 Ka). Nitrogen and carbon isotope data from skull collagen are discussed. The reason for the appearance of the walrus far from the modern sea shore was, presumably, the Rodionovo (Shklov, MIS 7) or boreal Sula (Mikulino, MIS 5e) marine transgression into the area of the latitudinal part of the Pechora River. The relatively good preservation of the bone testifies in favor of the Sula marine transgression. The Rodionovo age of the walrus can be assumed on the basis of the presence of shallow water marine deposits lying between two Middle Pleistocene moraines—Pechora (Dnieper, MIS 8) and Vychegda (Moscow, MIS 6).

Prista et al.: Considerations on the Portuguese Sirenia (Mammalia) fossil record.
Marine mammals of the Order Sirenia are good palaeoenvironmental proxies in paleontological studies, due to their specific feeding and habitat adaptations. Fossils of sirenians are rather common in the Portuguese Neogene, having been reported for nearly 100 years, although most findings correspond to rather fragmentary material, with no diagnostic value. Still, some nominal taxa of Sirenia were referred to in the literature, based either on identifiable fossils or on some undescribed material preserved in collections. Following revisions of published occurrences and new descriptions of previously unpublished material, a general review of the fossil sirenians of Portugal is presented, covering both the main geographic areas and the nominal taxa referred to in the literature. Several findings are corrected regarding the taxonomic classification, clearing the fossil record of Portugal, and making available for researchers the complete list of Sirenia findings and their geolocation in the Portuguese Neogene. An analysis of the Portuguese Sirenia fossil record allowed to solve problems associated to the previous published taxonomic works and increase the knowledge regarding the Lower Tagus Basin, Setubal peninsula, Alvalade Basin and Sirenia evolution in Portugal during the Lower and Middle Miocene. The Portuguese fossil record of sirenians shows evidence that these marine mammals were migrating towards south, which agrees with the European data, strengthening the present interpretation that sirenians took refuge in the Mediterranean and Gulf of Cadiz by the end of the Miocene, inhabiting these regions up to the beginning of the Northern Hemisphere Continental Glaciations by the end of the Piacenzian.

Ritsche and Hampe: Two exceptional Balaenomorpha (Cetacea: Mysticeti) from the Biemenhorst Subformation (middle/late Miocene) of Bocholt (W Münsterland, Germany) with a critical appraisal on the anatomy of the periotic bone.
Here we present two new Miocene basal balaenomorph mysticetes recognized by periotic bones with a unique combination of characters. The fossil periotics originated from former shallow marine sediments and were discovered in a suburban quarry of Bocholt in the German Münsterland holding a number of remains of a multispecies community of mysticetes. A detailed accompanying analysis of the internal and external structures of mysticete periotics resulted in a critical review and revision of two important characteristics of the pars cochlearis. The partition between the openings for the facial nerve and the vestibulocochlear nerve, named here ‘bony septum’ for baleen whales, has been developed so prominently during mysticete evolution that the crista transversa is no longer recognizable in derived forms, as it lies deep inside the inner ear canal and within the fundus of the internal acoustic meatus where it separates the cochlear nerve from the inferior vestibular nerve. We show that the designation of the opening for the superior vestibular nerve as ‘foramen singulare’ is misleading since this term describes the opening for the nerve to the posterior semicircular canal within the fundus of the internal acoustic meatus.

Skulls of living whales and dolphins (cetaceans) are telescoped—bones of the skull roof are overlapped by expanded facial bones and/or anteriorly extended occipital bones. Evolution of the underlying skull roof (calvarium), which lies between the telescoped regions, is relatively unstudied. We explore the evolution and development of the calvarium of toothed whales (odontocetes) by integrating fetal data with Oligocene odontocete fossils from North America, including eight neonatal and juvenile skulls of Olympicetus†. We identified two potential synapomorphies of crown Cetacea: contact of interparietals with frontals, and a single anterior median interparietal (AMI) element. Within Odontoceti, loss of contact between the parietals diagnoses the clade including Delphinida, Ziphiidae and Platanistidae (=Synrhina). Delphinida is characterized by a greatly enlarged interparietal. New fetal series of delphinoids reveal a consistent developmental pattern with three elements: the AMI and bilateral posterior interparietals (PIs). The PIs most resemble the medial interparietal elements of terrestrial artiodactyls, suggesting that the AMI of cetaceans could be a unique ossification. More broadly, the paucity of conserved anatomical relationships of the interparietals, as well as the fact that the elements often do not coalesce into a single bone, demonstrates that assessing homology of the interparietals across mammals remains challenging.
Comments: This study was fun to work on and while I collected data and made much of the figures, this was Rachel's brainchild and I largely took a backseat on this one - Jonathan has thought about telescoping for most of his career, and Rachel's an expert on the way skull bones grow and suture together. We basically kept finding interparietal bones in early odontocetes but they seemed to be different in layout and position: simocetids have three or four interparietals with a long leaf-shaped elemenet that forms along the midline and fuses to the parietals early. Waipatiids occasionally have a large, but wide interparietal; xenorophids have a plug-shaped interparietal that is forward-placed and frequently contacts the frontals and parietals. We named this median bone the AMI - anterior median interparietal - and tracked the occurrence of it and other interparietal bones amongst cetaceans, and found that the AMI might be a bone that is unique to Neoceti (we also reported it in Coronodon). These interparietal bones likely form as a result of increasing brain size in Odontoceti and a way to add some ossification centers on the top of the brain that can knit together with the parietals later on in growth - as the brain increases in size along the odontocete stem, the parietals are separated from one another longer during ontogeny, and in most odontocetes, completely separated throughout ontogeny.

Ruiz-Puerta et al.: Holocene deglaciation drove rapid genetic diversification of Atlantic walrus.
Rapid global warming is severely impacting Arctic ecosystems and is predicted to transform the abundance, distribution and genetic diversity of Arctic species, though these linkages are poorly understood. We address this gap in knowledge using palaeogenomics to examine how earlier periods of global warming influenced the genetic diversity of Atlantic walrus (Odobenus rosmarus rosmarus), a species closely associated with sea ice and shallow-water habitats. We analysed 82 ancient and historical Atlantic walrus mitochondrial genomes (mitogenomes), including now-extinct populations in Iceland and the Canadian Maritimes, to reconstruct the Atlantic walrus' response to Arctic deglaciation. Our results demonstrate that the phylogeography and genetic diversity of Atlantic walrus populations was initially shaped by the last glacial maximum (LGM), surviving in distinct glacial refugia, and subsequently expanding rapidly in multiple migration waves during the late Pleistocene and early Holocene. The timing of diversification and establishment of distinct populations corresponds closely with the chronology of the glacial retreat, pointing to a strong link between walrus phylogeography and sea ice. Our results indicate that accelerated ice loss in the modern Arctic may trigger further dispersal events, likely increasing the connectivity of northern stocks while isolating more southerly stocks putatively caught in small pockets of suitable habitat.

Rule et al.: Giant baleen whales emerged from a cold southern cradle.
Baleen whales (mysticetes) include the largest animals on the Earth. How they achieved such gigantic sizes remains debated, with previous research focusing primarily on when mysticetes became large, rather than where. Here, we describe an edentulous baleen whale fossil (21.12–16.39 mega annum (Ma)) from South Australia. With an estimated body length of 9 m, it is the largest mysticete from the Early Miocene. Analysing body size through time shows that ancient baleen whales from the Southern Hemisphere were larger than their northern counterparts. This pattern seemingly persists for much of the Cenozoic, even though southern specimens contribute only 19% to the global mysticete fossil record. Our findings contrast with previous ideas of a single abrupt shift towards larger size during the Plio-Pleistocene, which we here interpret as a glacially driven Northern Hemisphere phenomenon. Our results highlight the importance of incorporating Southern Hemisphere fossils into macroevolutionary patterns, especially in light of the high productivity of Southern Ocean environments.
Comments: the evolution of body size in mysticetes is one of my favorite debates in marine mammal paleontology - make sure to check out my blog post on this topic here.

Seppä et al.: Polar bear’s range dynamics and survival in the Holocene.
Polar bear (Ursus maritimus) is the apex predator of the Arctic, largely dependent on sea-ice. The expected disappearance of the ice cover of the Arctic seas by the mid 21st century is predicted to cause a dramatic decrease in the global range and population size of the species. To place this scenario against the backdrop of past distribution changes and their causes, we use a fossil dataset to investigate the polar bear's past distribution dynamics during the Late Glacial and the Holocene. Fossil results indicate that during the last deglaciation, polar bears were present at the southwestern margin of the Scandinavian Ice Sheet, surviving until the earliest Holocene. There are no Arctic polar bear findings from 8000–6000 years ago (8–6 ka), the Holocene's warmest period. However, fossils that date from 8-9 ka and 5–6 ka suggest that the species likely survived this period in cold refugia located near the East Siberian Sea, northern Greenland and the Canadian Archipelago. Polar bear range expansion is documented by an increase in fossils during the last 4000 years in tandem with cooling climate and expanding Arctic sea ice. The results document changes in polar bear's distribution in response to Late Glacial and Holocene Arctic temperature and sea ice trends.

In the southern region of the Baja California Sur state (BCS), Mexico has located Los Barriles sedimentary basin, which was part of the extension of the opening of the Gulf of California . Its evolution began in the late Miocene (8-7 Ma). The Trinidad Formation is found in this basin, with a late Miocene to Pliocene age. The marine sedimentary sequence is composed of mudstones, siltstones, and sandstones, interpreted as a shallow to deep water marine environment with gravity flows. In this area, in 1999, mysticete was collected at the El Cardonal locality on the La Higuera stream, consisting of a siltstone-sandstone concretion. In which there is an in situ tympanic bula and periotic complex with affinity to the genus Incakujira.

More than 23 extinct species and 10 extant species of the Balaenopteridae are known. Our knowledge of the family Balaenopteridae is increasing quickly, however, few fossil records support a circum-North Pacific distribution of balaenopterid genera and species. Because of limited preservations, most rorqual fossils reported from the western North Pacific can only be identified to the family level. A skull from the Shinazawa Formation (late Pliocene to Early Pleistocene) in Yamagata, Japan, is identified as aff. Balaenoptera bertae by possessing two diagnostic features of the species: large occipital condyles, and a posteriorly elongate postglenoid process. Combination of four more features also support that the specimen is a closely related to B. bertae. The specimen is probably a slightly older individual than the holotype of B. bertae, based on the estimated bizygomatic width and slightly longer posterior process of the tympanoperiotic. The first and only report of B. bertae was from the Pliocene Purisima Formation in California, USA. The specimen from Japan is incompletely preserved, but shows the occurrence of B. bertae in the western North Pacific for the first time, as many living balaenopterids are distributed across the North Pacific, such as Balaenoptera musculus, B. physalus, B. borealis, B. acutorostrata, and Megaptera novaeangliae.
Comments: This to me is one of the most personally gratifying papers to come out this year, even if it is one of the more humble papers in scope. This study reports a specimen of Balaenoptera bertae - a fossil I discovered and excavated when I was only 19 - from slightly younger rocks in Japan! This indicates that this species had a circum North Pacific distribution. Better yet, this paper was published by my friend Yoshi Tanaka, who I shared an office with during my 3.5 years in New Zealand! We disagreed on the thermostat in the office (he was perpetually warm, I was perpetually freezing) but we had a great time in that little office and got a ton of killer research done in there. Along with many, many discussions comparing Japan and the USA.

Tanaka et al.: A new specimen of cf. Isanacetus laticephalus (baleen whale) from the Oi Formation, Ichishi Group (late early Miocene) in Japan.
In the history of baleen whales, the early Miocene is one of not a well-known time. Many baleen whales from the Miocene are known as “cetotheres”. The “cetothere” Isanacetus laticephalus was a baleen whale from the early to middle Miocene that is only known from Japan. The holotype and paratype of I. laticephalus are well-preserved, and the age of the species I. laticephalus makes it one of the earlier basal plicogulans; however, its phylogenetic position is unclear. Here, a new cranium including the periotic from an early Miocene deposit (about 18.5 to 17.0 Ma) of the Oi Formation, Ichishi Group in Mie, Japan is reported as cf. I. laticephalus. The specimen displays a combination of four periotic features: (i) a large lateral tuberosity; (ii) a rectangular pars cochlearis; (iii) a robust neck of the posterior process, and (iv) a cerebral aperture of the facial canal that is smaller than that of the dorsal vestibular area. This combination of features could be diagnostic for I. laticephalus, among basal plicogulans from the early and middle Miocene. Adding more referred specimens will expand the diagnoses of I. laticephalus, and firmly support phylogenetic trees with this species.

Torcarescu: Cetotheriidae (Cetacea, Mysticeti) from the Collections of the National Geological Museum, Bucharest (Romania.Cetacean fossil remains attributed to Cetotheriidae are certainly not a novelty in the fossil record of the Carpathian Foreland or Foredeep, several sites being mentioned; however there remains much material in institutional collections that can still add to the number of fossiliferous localities where cetotheriids can be found. The aim of this paper is to bring to attention new material from fossiliferous localities from the western Dacian Basin (Gorj and Vâlcea Counties), from the collections of the National Geological Museum. The specimens are described, identified based on comparisons with similar specimens from published literature and a discussion about the species Cetotherium priscum is provided.

Tosetto et al.: Cranial Material of Long-Snouted Dolphins (Cetacea, Odontoceti, Eurhinodelphinidae) from the Early Miocene of Rosignano Monferrato, Piedmont (NW Italy): Anatomy, Paleoneurology, Phylogenetic Relationships and Paleobiogeography.
We provide a new study of previously published eurhinodelphinid materials from the early Miocene of Piedmont (NW Italy) based on a new preparation of the fossil specimens. We studied specimens previously assigned to Tursiops miocaenus and Dalpiazella sp. and provide new anatomical data on the eurhinodelphinid skull and ear bones. In particular, we suggest that a skull that was previously assigned to Tursiops miocaenus must be reassigned to Ziphiodelphis sigmoideus (Cetacea, Odontoceti, Eurhinodelphinidae) based on new comparisons of the squamosal. This finding enabled us to provide new anatomical information on the ear bone anatomy of Z. sigmoideus that was previously unknown. The material originally assigned to Tursiops miocaenus is currently lost. For this reason and due to the fact that the partial illustration of this species by Portis does not allow us to find diagnostic characters for this species, we decided that Tursiops miocaenus is a nomen dubium. Analysis of additional isolated teeth previously assigned to Tursiops miocaenus led to the conclusion that these specimens represent Odontoceti incertae sedis. We performed a new phylogenetic analysis by adding newly discovered character states to a previous dataset and a paleobiogeographic analysis of Eurhinodelphinidae. We found two monophyletic clades within this family. The paleobiogeographic pattern found by the present work suggests the existence of North Atlantic and Mediterranean clades with some species distributed among both basins. We analyzed the virtual endocast of Ziphiodelphis sigmoideus and found that it resembles that of Schizodelphis in several respects, suggesting that some of the more derived characters of the odontocete brain were still absent in these early Miocene eurhinodelphinids.

Tsai: In search of the origin of crown Mysticeti.
Recent research on mysticete fossils from the Late Eocene and Oligocene has revolutionised our understanding of the diversity and evolutionary scenarios for early baleen whales. For example, aetiocetids are a possible, though controversial, lineage that bridges the gap between the toothed and baleen-bearing mysticetes, and eomysticetids show a further transitional step towards the baleen-bearing status, with the presence of non-functional dentition in adults. However, information about the origin of crown mysticetes, including the most recent common ancestor of all extant lineages and its descendants, is critical to further understanding the evolution of baleen whales. The phylogenetic positions of the Oligocene Toipahautea, Whakakai, Horopeta, and Mauicetus from New Zealand remain unresolved and problematic, but all four genera show a close relationship with crown mysticetes. The original and subsequent cladistic analyses have consistently revealed a sister relationship between the Toipahautea-to-Mauicetus grade and crown mysticetes, and Horopeta has been placed close to the cetotheriids within the crown group. This review aims to stimulate more research on this topic by elucidating the origin of crown mysticetes, which likely experienced a poorly known radiation event during the Oligocene that established the modern lineages.

Van Vliet et al.: An archaeocete vertebra re-examined: indications for a small-sized species of Pachycetus from Spain, Europe.
A cetacean vertebra from presumably Bartonian strata near Taradell, Northeastern Spain, described by Pilleri in 1989, is here re-examined. Also a description of the local stratigraphy is given. Notwithstanding the rather severe damage of the vertebral centrum, it shows the same characteristics as vertebrae ascribed to Pachycetus (‘Platyosphys’; ‘Basilotritus’) sp. from, amongst others, Ukraine, Germany and Belgium. The characteristics are: the combined presence of a compact, circumferential multi-layered cortex, the pock-marked surface of the vertebral centrum, the pachyostotic pedicles of the neural arch and finally the probable elongation of the vertebral centrum, the transverse processes and the pedicles of the neural arch. The compactness of the midpart of the vertebral centrum has been measured and is compared with that of other taxa. The find extends the geographic distribution of this genus to Southwestern Europe. The vertebra appears to belong to a small species of Pachycetus, which was until now not known with certainty from Europe, although some similar-sized vertebrae from the Southern Bight of the North Sea have recently been described.

Velez-Juarbe: New heterodont odontocetes from the Oligocene Pysht Formation in Washington State, U.S.A., and a reevaluation of Simocetidae (Cetacea, Odontoceti).
Odontocetes first appeared in the fossil record by the early Oligocene, and their early evolutionary history can provide clues as to how some of their unique adaptations, such as echolocation, evolved. Here, three new specimens from the early to late Oligocene Pysht Formation are described further increasing our understanding of the richness and diversity of early odontocetes, particularly for the North Pacific. Phylogenetic analysis shows that the new specimens are part of a more inclusive, redefined Simocetidae, which now includes Simocetus rayi, Olympicetus sp. 1, Olympicetus avitus, O. thalassodon sp. nov., and a large unnamed taxon (Simocetidae gen. et sp. A), all part of a North Pacific clade that represents one of the earliest diverging groups of odontocetes. Amongst these, Olympicetus thalassodon sp. nov. represents one of the best known simocetids, offering new information on the cranial and dental morphology of early odontocetes. Furthermore, the inclusion of CCNHM 1000, here considered to represent a neonate of Olympicetus sp., as part of the Simocetidae, suggests that members of this group may not have had the capability of ultrasonic hearing, at least during their early ontogenetic stages. Based on the new specimens, the dentition of simocetids is interpreted as being plesiomorphic, with a tooth count more akin to that of basilosaurids and early toothed mysticetes, while other features of the skull and hyoid suggest various forms of prey acquisition, including raptorial or combined feeding in Olympicetus spp., and suction feeding in Simocetus. Finally, body size estimates show that small to moderately large taxa are present in Simocetidae, with the largest taxon represented by Simocetidae gen. et sp. A with an estimated body length of 3 m, which places it as the largest known simocetid, and amongst the largest Oligocene odontocetes. The new specimens described here add to a growing list of Oligocene marine tetrapods from the North Pacific, further promoting faunistic comparisons across other contemporaneous and younger assemblages, that will allow for an improved understanding of the evolution of marine faunas in the region.
Comments: Between this and the Xenorophus monograph, it's been a banner year for stem odontocetes! This is a very important study, an in-depth followup to the Jorge's 2017 paper naming Olympicetus. This monograph reports a new species of Olympicetus, Olympicetus thalassodon, along with Olympicetus sp. 1 and a fourth Olympicetus-like simocetid. The phylogenetic analysis in this study firmly places Olympicetus into a clade with CCNHM 1000 (from Racicot et al., 2019) and Simocetus - and these taxa are formally identified as belonging to the Simocetidae. Which is really convenient, because I've been lazily calling Olympicetus a simocetid for years.

The Palaeontology-Stratigraphy Museum is hosted by the Directorate for Heritage of the “Babeș-Bolyaiˮ University from Cluj-Napoca. Its collections were founded in the 19th century, then gradually enriched through donations of the professors and students, or by exchanges with other museums. They were initially used for teaching and research purposes and are nowadays available for the public as well. The sirenian fossils in the museum’s collection, even though are not many, represent an important heritage, a valuable tool in illustrating the Paleogene marine fauna of Romania and the worldwide fossils of these marine mammals. Today, the museum hosts 39 pieces of sirenian fossils. The aim of this article is an inventory of these fossils and the revision of the data referring to each piece. This paper includes the following data: the material, inventory numbers, the name of the locality where the fossil originated from, the stratigraphic unit, the geological age, the name of donor when available and, in few cases, measurements and short descriptions of the pieces, when necessary.

Variations in the shape and size of teeth have been associated with changes in enamel ultrastructure across odontocetes. Characterizing these features in extinct taxa can elucidate their functional morphology and feeding strategy, while also shedding light into macroevolutionary patterns during the evolutionary history of cetaceans. This study aimed to (1) describe the enamel and dentine ultrastructure of the Early Miocene odontocetes Notocetus vanbenedeni and Phoberodon arctirostris from Patagonia (Argentina) and (2) quantify tooth and enamel ultrastructure morphological disparity among odontocetes. Enamel was predominantly prismatic, thin in the anterior tooth of N. vanbenedeni and P. arctirostris; whilst thick on the posterior tooth of N. vanbenedeni. Together with skull morphology, data suggests a raptorial feeding strategy for P. arctirostris and a combination suction feeding method for N. vanbenedeni. Statistical analyses supported these inferences, indicating that enamel characters are useful for paleoecological research. Morphological disparity analyses showed that extant odontocetes occupy a larger morphospace and have more disparate morphologies, whilst extinct odontocetes were more similar among each other than with the extant group. There was no clear phylogenetic-based grouping, suggesting that tooth and enamel ultrastructure disparity were mainly driven by ecological pressures. These results highlight enamel ultrastructure as a source for broader-scale paleoecological studies in cetaceans.

Viglino et al.: Aquatic mammal fossils in Latin America – a review of records, advances and
challenges in research in the last 30 years.
Records of aquatic mammal fossils (e.g. cetaceans, pinnipeds, sirenians, mustelids, and desmostylians) from Latin America (Mexico to Tierra del Fuego, including Antartica) span since the mid-1800s. Aquatic mammal fossils received little attention from the scientific community, with most of the first studies conducted by Northern Hemisphere researchers. Over the last 30 years, paleontological research in Latin America has increased considerably, with descriptions of several new species and revisions of published original records. The Latin American fossil record of marine mammals spans from the Eocene to the Pleistocene, with formations and specimens of global significance. All three main groups of cetaceans are represented in the continent (Archaeoceti, Mysticeti, and Odontoceti). Pinnipedia are represented by the families Otariidae and Phocidae, with records starting in the Middle Miocene. Both living families of Sirenia (Trichechidae and Dugongidae) are recorded. While less common, but still relevant, records of desmostylians and mustelids are known from Oligocene and Miocene deposits. This review provides a summary of the aquatic mammals known to date, with a special focus on the advances and developments of the last 30 years, since Cozzuol’s (1996) review of the South American fossil record. An up-to-date complete list of species based on the literature and unpublished data is also provided. The study also provides future directions for paleontological research in Latin America, and discusses the challenges and opportunities in the field, including the emergence of a strong new generation of Latin American researchers, many of whom are women.
Comments: I've skimmed this paper, and it is one I really need to sit down and read again more carefully: I have a feeling that this is one of the most important papers of the year. This new genre of papers - reflections on our own discipline and studies of ethics - is a welcome new direction and I often feel that these are more interesting than a steak-and-potatoes description of a new species.

Voss et al.: New findings of Prototherium ausetanum (Mammalia, Pan-Sirenia) from paving stones in Girona (Catalonia, Spain)?
Taxonomic and morphological approaches on Eocene sirenians from Catalonia (Spain) benefit from a newly discovered specimen found in a quite unusual locality, the pedestrian zone in the city of Girona. Two fossil-bearing limestone slabs from middle Eocene (Bartonian) layers of a quarry in the wider surrounding area north-west of Barcelona, were CT-scanned in the Clínica Girona to enhance more detailed investigations. Post-processing of the scans and, as far as possible, 3D-reconstruction of the preserved elements in the slabs was performed at Museum für Naturkunde Berlin. Thereby, a skull of a Dugong specimen was used as a reference point. Based on the combined analysis of macroscopic and CT-data, the specimen most likely represents Prototherium ausetanum Balaguer & Alba, 2016 and complements the available information of the holotype and hitherto only known specimen of that species. The Girona specimen is an adult, but small individual that corroborates P. ausetanum as a generally small-sized species compared to other known Prototherium taxa.