Oi,
it's been a long time since I've done this. We're five months late here, but
let's just say 2019 was not a great year for me professionally and it's taken
me about a year to mentally catch up, accept the new normal, and re-motivate me
to do research. Have you ever seen Office Space? I was Peter Gibbons for quite
a while (still am, and my students love it). Anyway, I'm going to get back into
this blogging thing. Truth be told, winter was very busy for me, and so I
genuinely did not have any time to write this post on time - all of December
was spent working on getting a cast of a whale skeleton repaired and painted
and ready for mounting. I reviewed like 16 manuscripts in 2019, and didn't
submit a single one of my own! I finally, for the first time in my career,
declined a few requests to keep my stress levels down. I taught my first giant
lecture with about 100 students - which was not a whole lot harder than our
typical 50 student lectures (more emails, though). And, now we're stuck in
quarantine - and I've been catching up on various things in a big way, so
without further ado, brought to you by Covid19, the very belated 2019 in marine
mammal paleontology post!

Albright
et al.: Cenozoic vertebrate biostratigraphy of South Carolina, U.S.A., and additions to the fauna.
Bulletin of the Florida Museum of Natural History.
Link:
https://digitalcommons.unf.edu/facultyshowcase/2020/Showcase/1/
Abstract:
Study of vertebrate fossils from the South Carolina Coastal Plain played a
significant role in the early history of vertebrate paleontology as a
scientific discipline in North America. However, a clear understanding
of the state’s vertebrate biostratigraphy has been greatly hindered by the
paucity of well-exposed fossil-bearing stratigraphic sections and a complicated
subsurface stratigraphy. Most units, particularly those of Neogene and
Quaternary age, exist as thin veneers of marine or estuarine sediments that
typically occur as infilled topographic lows or erosional remnants as
determined primarily from borehole stratigraphy. Hence, lateral continuity can
be difficult to confirm over broad geographic areas often resulting in
confusion insofar as vertebrate fossil provenance is concerned. The evaluation
of vertebrate fossils from the South Carolina Coastal Plain presented here, and
of the geologic units from which they originated, or are thought to have
originated, results in the first modern biostratigraphic framework for the
known Cenozoic record of the state. Results provide (1) updated correlations of
many units to the most recent, astronomically-tuned marine oxygen isotope
stages; (2) the first viverravid from the Atlantic Coastal Plain, Didymictis
proteus; (3) new information on the timing of certain Neotropical immigrants
into the Southeast during the Great American Biotic Interchange, including the
oldest record of Erethizon in this region and the oldest records of capybara in
the USA; (4) the possible oldest record of the microtine rodent Allophaiomys
pliocaenicus in the USA; (5) new details on the unit of origin for the type
specimen of the archaeocete Dorudon serratus; and (6) new details on the
ontogeny of the early odontocete Agorophius pygmaeus. New taxonomic records
from South Carolina include Glyptotherium texanum, Holmesina
floridanus, Ondatra idahoensis, Erethizon ?bathygnathum, Leopardus amnicola,
Miracinonyx ?trumani, Canis lepophagus, Canis latrans, Phocanella pumila,
Callophoca obscura, Monatherium sp., Anchippus texanus, Subhyracodon mitis,
Aphelops ?malacorhinus, Teleoceras ?guymonense, and Perchoerus sp. New
specimens of previously recorded taxa are also noted.
Comments:
This behemoth of a paper has a LOT in it, and once I have barely had time to
scratch the surface of - but discusses the South Carolina marine vertebrate fossil
record in detail, including all sorts of obscure historical references I wasn't
aware of. Also, it's worth noting that because South Carolina is so anomalously
fossiliferous, and there are also no cliffs, that the history of paleontology
here is dominated by tons and tons and tons of very neat but context-less ex
situ fossils with uncertain ages and disagreements over provenance.
Bianucci
et al: A new Monodontidae (Cetacea, Delphinoidea) from the lower Pliocene of
Italy supports a warm-water origin for narwhals and white whales. Journal of
Vertebrate Paleontology.
Link:
https://www.tandfonline.com/doi/abs/10.1080/02724634.2019.1645148?journalCode=ujvp20
Abstract:
A new taxon of monodontid cetacean, Casatia thermophila, gen. et
sp. nov., is here described on the basis of a partial skull from lower Pliocene
(5.1–4.5 Ma) marginal-marine deposits of Tuscany (central Italy). This new taxon belongs to
Monodontidae based on the presence of a medial exposure of the maxillae
anterior and lateral to the external bony nares; it mainly differs from all
other named monodontids by the presence of a median depression of the
premaxillae anterior to the premaxillary sac fossae and by a medial margin of
the premaxillary-maxillary suture that does not parallel the anterolateral
profile of the external bony nares. Our phylogenetic analysis, the first
including all taxa of Monodontidae, recovers Casatia as a
crown monodontid, more closely related to Delphinapterus than
to Monodon and sister group of an unnamed taxon from the North
Sea. The holotype of Casatia represents the first and only
fossil monodontid from the Mediterranean Basin. Taking its place beside
abundant fossils of strongly thermophilic marine vertebrates, such as the bull
shark Carcharhinus leucas, the tiger shark Galeocerdo
cuvier, and the extinct sirenian Metaxytherium subapenninum, Casatia
thermophila represents the strongest evidence supporting the
hypothesis that monodontids once thrived in low-latitude, warm-water habitats.
On the basis of our phylogenetic reconstruction, early relatives of the extant
monodontids might have adapted independently to the high-latitude, cold-water
environments they currently master. The definitive disappearance of the Neogene
thermophilic monodontids could be attributed to the cooling episode that
accompanied the onset of long-term Northern Hemisphere glaciation around 3 Ma.
Comments:
This paper is sort of a response to the 2018 paper by Ichishima et al. on
Haborodelphis, a beluga-like monodontid from the Pliocene of Hokkaido. Previously
reported temperate to subtropical latitude monodontids (e.g. Denebola,
Bohaskaia) suggested that the earliest belugas were actually warm water
cetaceans - but Ichishima et al. found a cold water invertebrate assemblage,
and indicated that since there are some quite cold (albeit brief) periods in
the Pliocene, greater care was needed to demonstrate paleoclimate aside from
"ahhh, yeah it's from Mexico so it's gotta be warm". Casatia
thermophila (great species name!) was recovered with a nice assemblage of warm
water taxa, in the Mediterranean! Given the timing and incomplete preservation of Casatia I do wonder if
Bianucci et al. saw the Ichishima paper and thought "well now is the
perfect time to get this thing published" and wrote it up. Fascinating
little back and forth.

Bianucci
et al. Rise of the titans: baleen whales became giants earlier than thought.
Biology Letters.
Link: https://royalsocietypublishing.org/doi/10.1098/rsbl.2019.0175
Abstract:
Baleen whales (Mysticeti) are major ecosystem engineers, thanks to their
enormous size and bulk filter feeding strategy. Their signature gigantism is
thought to be a relatively recent phenomenon, resulting from a Plio-Pleistocene
mode shift in their body size evolution. Here, we report the largest whale
fossil ever described: an Early Pleistocene (1.5–1.25 Ma) blue whale from Italy
with an estimated body length of up to 26 m. Macroevolutionary modelling taking
into account this specimen, as well as additional material from the Miocene of
Peru, reveals that the proposed mode shift occurred either somewhat earlier, or
perhaps not at all. Large-sized mysticetes comparable to most extant species
have existed since at least the Late Miocene, suggesting a long-term impact on
global marine ecosystems.
Comment:
This paper is a response of sorts to earlier studies, chiefly Slater et al.
2017, which proposed that baleen whales did not become gigantic (e.g.
over 15 meters) until the last 2-3 million years - the analysis was based on
skull size and found that gigantism evolved independently and late in balaenids
and balaenopteroids. How late, nobody was sure, because most Pliocene fossil
mysticetes are still on the small side and our Pleistocene record is terrible.
I always thought the Slater hypothesis was correct, based on the lack of large
mysticete *crania*. This new study reported a nearly full size fossil blue
whale skull from the middle Pleistocene of Italy, and a few gigantic mysticetes
from the latest Miocene of Peru (Pisco Fm., of course) and it pulled the onset
of gigantism back a few million years - same overall pattern, just slightly
earlier. Now: I think a better way around this, to get a better sample size, is
to get a regression for body length based on vertebral size (which, based on my
last project, is eminently possible) and measure hundreds and hundreds of
isolated vertebrae. Granted, vertebrae are non-diagnostic, so they will be
devoid of phylogenetic context, but may show a very different pattern as I have
seen gigantic vertebrae in cliffs, backyards, and museum collections from late
Miocene and Pliocene localities, and I believe there is certainly collecting
bias against large mysticetes (too big to excavate).

Bisconti
et al. A new balaenopterid whale from the late Miocene of the Southern North
Sea Basin
and the evolution of balaenopterid diversity (Cetacea, Mysticeti). PeerJ.
Link:
https://peerj.com/articles/6915/
Abstract:
Balaenopterid mysticetes represent the most successful family-rank group of
this clade. Their evolutionary history is characterized by a rich fossil record
but the origin of the living genera is still largely not understood. Recent
discoveries in the southern border of the North Sea revealed a number of well
preserved fossil balaenopterid whales that may help resolving this problem. In
particular, skull NMR 14035 shares morphological characters with the living
humpback whale, Megaptera novaeangliae and, for this reason,
its characteristics are investigated here. The comparative anatomical analysis
of the new specimen formed the basis of a new phylogenetic analysis of the Mysticeti
based on a matrix including 350 morphological character states scored for 82
Operational Taxonomic Units. The stratigraphic age of the specimen was
determined based on the analysis of the dinocyst assemblage recovered in the
associated sediment. We assessed clade diversity in Balaenopteridae by counting
the numbers of clades in given time intervals and then plotted the results. Nehalaennia
devossi n. gen. et sp. is described for the first time from the late
Tortonian (8.7–8.1 Ma) of the Westerschelde (The Netherlands). This new taxon
belongs to Balaenopteridae and shows a surprisingly high number of advanced
characters in the skull morphology. Nehalaennia devossi is
compared to a large sample of balaenopterid mysticetes and a phylogenetic
analysis placed it as the sister group of a clade including the genus Archaebalaenoptera.
The inclusion of this fossil allowed to propose a phylogenetic hypothesis for
Balaenopteridae in which (1) Eschrichtiidae (gray whales) represents a family
of its own, (2) Balaenopteridae + Eschrichtiidae form a monophyletic group
(superfamily Balaenopteroidea), (3) Cetotheriidae is the sister group of
Balaenopteroidea, (4) living Balaenoptera species form a
monophyletic group and (5) living M. novaeangliae is the
sister group of Balaenoptera. Our work reveals a complex
phylogenetic history of Balaenopteridae and N. devossi informs
us about the early morphological transformations in this family. Over time,
this family experienced a number of diversity pulses suggesting that true
evolutionary radiations had taken place. The paleoecological drivers of these
pulses are then investigated.
Comments:
An interesting new study reporting the new genus and species Nehaelennia
devossi - an Archaebalaenoptera-like early rorqual. When I started working on
fossil baleen whales, we were in a state of 'taxonomic paralysis' because of
good ole P.J. Van Beneden, who, bless him, named a lot of fossil species of
mysticetes from Belgium in the late 19th century - based on chimaeric
assemblages of non-associated fossils that he arranged to fit pre-imagined
'archetypes' (e.g. the modern Atlantic fauna has a small minke whale, so all
these small bones go to the Pliocene version of the minke whale and surely all
go together... right?) and then like, went ahead and fucking renamed a bunch of
his own species and genera! Anyway, nobody had the gall to start fixing the
problem until 2010 when Bosselaers and Post shit-canned all of his
balaenopterids as nomina dubia with the possible exception of "Balaenoptera" borealina.
Since then a number of new discoveries of Belgian and Dutch mysticetes have
finally been able to be studied without being mired in the taxonomic web of Van
Beneden. This new paper says a lot of things, including a very different
placement of my own species Balaenoptera bertae which I very much doubt.
Regardless, the new specimen is nice.

Boessenecker
and Boessenecker. Paleontology of the "Ashley Phosphate Beds"
of Charleston: insights from Northbridge Park, Charleston, South Carolina. GSA Field Guidebooks.
Link:
https://pubs.geoscienceworld.org/books/book/2152/chapter/120859059/Paleontology-of-the-Ashley-Phosphate-Beds-of
Abstract:
A man-made deposit at Northbridge Park near Charleston, South Carolina, consists of phosphatic
nodules, fossils, and mud dredged from the bottom of the Ashley River; nodules and fossils lay strewn
across the banks of the river. This artificial deposit is likely representative
of deposits mined extensively in the late nineteenth century and widely
referred to as the “Ashley Phosphate Beds.” Many of the taxa discovered at
Northbridge Park were historically reported from the phosphate beds, and
include sharks, rays, bony fish, sea turtles, giant birds, whales, dolphins,
sea cows, and land mammals. Some of these bear adhering matrix indicating
origin from the Oligocene Ashley Formation. Others lack matrix but have short
geochronologic ranges and are derived from the Ashley Formation, Lower Miocene
Marks Head Formation, Lower Pliocene Goose Creek Limestone, and Pleistocene
Wando Formation.
Comments:
This paper hilariously started out as a blog post (elsewhere on this very blog) and as the result of many
recreational fossil collecting trips to a fossil site near my old apartment.
This was the first fossil site I was able to visit now that quarantine closures
are easing. GSA publications are notoriously hard to get a hold of, so email me
if you want a pdf. This was my only paper in 2019! Coauthored with my lovely
wife and coworker.
Bosio.
Integrated chronostratigraphy and taphonomic studies in Miocene sedimentary
successions of the East Pisco Basin (Peru). Plinius.
Link:
http://www.socminpet.it/Plinius2019/bosio.pdf
Abstract:
(no abstract, so here's the introduction) Along the western side of the Ica
River (Ica Desert, Peru), the Miocene Chilcatay and Pisco formations of the
East Pisco Basin crop out. These units are characterized by an extraordinarily
preserved marine vertebrate fossil assemblage, especially cetaceans. This work
has the purpose of making chronostratigraphic and taphonomic studies on these
formations, cropping out in the Ica River valley. The Pisco Basin, in southern Peru, is one of the fault-bounded
Cenozoic sedimentary basins along the Peruvian coast, located at 14°30’ S of
latitude. Along the coast, the most easterly Outer Shelf High separates the
forearc Pisco Basin into an offshore and an
onshore portion, referred to as the West Pisco Basin, still submerged, and East Pisco Basin (Fig. 1a). The sedimentary
infill of the East Pisco Basin has been described by Dunbar et al. (1990) and
DeVries (1998), and consists of five lithostratigraphic units: the Eocene
Caballas Formation, the
middle to upper Eocene Paracas Formation (subdivided in Los Choros and Yumaque
members), the uppermost Eocene-lower Oligocene Otuma Formation, the uppermost
Oligocene to lower Miocene Chilcatay Formation, and the upper Miocene to
Pliocene Pisco Formation (Fig. 1b) (e.g., Dunbar et al., 1990; DeVries, 1998;
DeVries & Jud, 2018).
Comments:
I think there will be a longer followup paper later - I always look forward to
taphonomic research!
Buono
et al. Eocene Antarctica: a window into the earliest history of modern whales.
Advances in Polar Science.
Link:
http://www.aps-polar.org/paper/2019/30/03/A190617000001
Abstract:
The Eocene–Oligocene Southern Ocean is thought to have played a major role in
cetacean evolution. Yet, fossils from its heart—Antarctica—are rare, and come almost
exclusively from the Eocene La Meseta and Submeseta formations of Marambio
(Seymour) Island. Here, we provide a summary and update of this crucial fossil
assemblage, and discuss its relevance in the broader context of cetacean
evolution. To date, Eocene specimens from Antarctica include basilosaurids, a
group of archaic stem cetaceans that had already fully adapted to life in
water; and the archaic toothed mysticete Llanocetus, the second oldest crown
cetacean on record (ca. 34 Ma). This Eocene co-occurrence of stem and crown
cetaceans is highly unusual, and otherwise only observed in Peru. Though related, at least some
of the Antarctic species appear to be different from, and notably larger than
their Peruvian counterparts, suggesting an early differentiation of the high
latitude cetacean fauna.
Comments:
This interesting new paper is a summary of recent papers on basilosaurids and
the toothed mysticete Llanocetus from the Priabonian La Meseta Formation of
Seymour Island, Antarctica. This unit is an important window into mid-Cenozoic marine vertebrate
evolution in the southern ocean. This paper does report some interesting new
specimens including a beautifully preserved mysticete pelvis *probably* (but
not certainly) belonging to Llanocetus.
Churchill
and Uhen. Taxonomic implications of morphometric analysis of earless seal limb
bones. Acta Palaeontologica Polonica.
Link:
https://www.app.pan.pl/article/item/app006072019.html
Abstract:
Fossil Phocidae (earless seals) are mostly known from isolated postcranial
material, forcing researchers to rely upon humeri and femora for the diagnosis
of taxa and reconstruction of phylogeny. However, the utility of these elements
has never been rigorously tested. Here, we provide the first quantitative
analysis of morphometric data from the humerus and femur, incorporating
measurement data from all extant genera as well as several fossil taxa.
Principle components analysis (PCA) found that genera clustered together on PC1
and PC2, although there was poor segregation of taxa and extensive overlap with
genera in adjacent regions of the morphospace. Discriminant function analysis (DFA)
was able to sort fossil taxa into different subfamilies, but performed poorly
at lower taxonomic levels. A preliminary review of phylogenetic characters
found that while some characters performed well at distinguishing different
subfamilies, many characters were poorly defined and not quantified, possessed
greater individual variation than past studies suggested, or were more variable
in fossil taxa. Our analyses suggest that the utility of isolated humeri and
femora for diagnosis of new taxa has been greatly exaggerated, and that extreme
caution should be applied to interpretations of taxonomy of fossil material
based on isolated elements. Future research should instead focus on study of
associated skeletons and cranial material. A thorough revision of fossil phocid
taxonomy is needed, and many described taxa are likely to be nomina dubia and
of limited use in phylogenetic analysis.
Comments:
This sorely needed paper is a first stab at actually resolving the hopelessly
confused study of fossil seal postcranial bones. Lots of noise has been made
about limb bones being diagnostic and paleoecologically informative, with
non-associated bones being jumbled together into discrete
"ecomorphotypes". This paper critically evaluates these claims and finds
that there is not much statistical support for the diagnostic value of isolated
limb bones.
Collareta
et al. A new kogiid sperm whale from northern Italy supports psychrospheric
conditions in the early Pliocene Mediterranean Sea. Acta Palaeontologica
Polonica.
Link:
https://app.pan.pl/article/item/app005782018.html
Abstract:
Among living cetaceans, dwarf and pygmy sperm whales (Kogia) are the
only members of the family Kogiidae, regarded as diminutive and elusive
relatives of the great sperm whale Physeter. Kogiids are known as
fossils by several skulls, teeth, and ear bones from Neogene deposits of the
Northern Hemisphere and Peru. We report on a fossil kogiid
specimen collected at Sant’Andrea Bagni (northern Italy) from Zanclean marine mudstone;
these deposits also yielded a rich deep-water elasmobranch assemblage depicting
the presence of Atlantic-derived psychrospheric waters. The kogiid specimen,
consisting of a partial cranium, one detached tooth, one vertebra, and one
fragmentary rib, is here referred to Pliokogia apenninica gen.
et sp. nov. Pliokogia is mostly characterised by a long and
dorsally flattened rostrum and by the presence of two well-distinct fossae on
the right side of the supracranial basin, including an elongated peripheral maxillary
fossa on the posterior portion of the right maxilla. Our phylogenetic analysis
recovers Pliokogia as a member of the subfamily Kogiinae,
which includes Kogia, Koristocetus, Nanokogia,
and Praekogia. A low temporal fossa and the absence of dental enamel
suggest that, like extant Kogia, Pliokogia was a suction
feeder. Since living kogiids do not inhabit the Mediterranean waters, and
considering that they feed on deep-water prey in open-sea areas, the
association of Pliokogia with a psychrospheric elasmobranch
assemblage with Atlantic affinities is noteworthy. Indeed, in early Pliocene
times, the Gibraltar connection was controlled by estuarine dynamics, thus allowing the
entrance of deep-water organisms (including the putative prey of Pliokogia)
in the Mediterranean Basin. The subsequent abandonment of
the Mediterranean Sea by kogiids might therefore be related to the definitive
establishment of the present-day antiestuarine circulation at Gibraltar, which
likely led to a limited deep nutrient supply and resulted in the strong
depletion of most Mediterranean deep-water ecosystems.

Cortes
et al. Shark‑cetacean trophic interactions during the late Pliocene
in the Central Eastern Pacific (Panama). Palaeontologia Electronica.
Link:
https://palaeo-electronica.org/content/2019/2652-pliocene-whale-from-panama
Abstract:
We provide a description of the remains of a fossil whale from western Panama. The record consists of
appendicular remains of a mysticete, which has been assigned to
Balaenopteridae. These remains, found in the sediments of the late Pliocene
Burica Formation, represent the first record of a marine mammal in the Neogene
sedimentary succession of the Burica Peninsula. Two different types of shark
bite marks, serrated and deep-unserrated, found on the radius and phalanges
suggest scavenging by at least two white shark (Carcharodon)
individuals. The deep, unserrated marks were possibly caused by continual
biting by sharks. Both the morphology of the shark bite marks and their
relative location on the whale limb bones constitute evidence of shark-cetacean
trophic interaction. Although the specimen lacks diagnostic features that would
allow a species-level identification, it does provide new information on the
vertebrate fauna of a very poorly prospected Central Eastern Pacific exposure,
thus opening an opportunity for exploring the marine fauna during a critical
episode in Earth history, the Plio-Pleistocene transition.

Cuitino
et al. Factors affecting the preservation and distribution of cetaceans in the
lower Miocene Gaiman Formation of Patagonia, Argentina. Palaeogeography,
Palaeoclimatology, Palaeoecology.
Link:
https://www.sciencedirect.com/science/article/abs/pii/S0031018218306904
Abstract:
Factors affecting cetacean preservation and distribution in ancient sedimentary
successions are not well understood. Actualistic studies have focused on
coastal and deep marine examples, and little is known about taphonomic
processes occurring within shelf environments. In this paper, we integrate
sedimentological, taphonomic and palaeontological data for the cetacean-bearing
lower Miocene Gaiman Formation from Patagonia, Argentina, to analyse which factors
affected the distribution and preservation of cetaceans in several localities
of the Southwestern Atlantic. Facies analysis shows that the formation
represents a transgressive-regressive stratigraphic cycle, with
palaeoenvironments including coastal, storm-dominated shoreface, inner shelf
embayment and open inner shelf. Cetacean remains show preservation styles
varying from isolated elements to articulated specimens, with nearly all
remains restricted to both embayment and open marine inner shelf settings. Two
fossil cetacean assemblages are recognized: Assemblage A comprises small-sized
odontocetes dominated by Platanistoidea, preserved mostly in inner shelf
embayment deposits; and Assemblage B comprises large-sized odontocetes and
mysticetes, dominated by Physeteroidea and Balaenopteroidea, preserved mostly
in open inner shelf deposits. Cetacean preservation style is controlled by a
combination of palaeoenvironmental conditions, such as energy and sedimentation
rate, and by biological factors, such as cetacean body plan and its impact on
carcass buoyancy. Ecological factors may also explain the stratigraphic
distribution of cetaceans between coastal and oceanic assemblages. This novel
integrative and multidisciplinary analyses improves our knowledge of cetacean
taphonomy in shelf environments.
Comments:
This study claims to show evidence of habitat preference in ancient cetacean
assemblages - I am skeptical owing to the process of bloat and float, and
wonder what a larger sample of additional assemblages would show. Nevertheless,
it is an interesting taphonomic study and food for thought.
Davydenko
et al. The earliest record of a marine mammal (Cetacea: Basilosauridae) from
the Eocene of the Amazonia. Journal of Vertebrate Paleontology.
Link:
https://www.tandfonline.com/doi/full/10.1080/02724634.2018.1549060
Abstract:
The Amazon region is known by its rich Cenozoic mammalian diversity; however,
fully aquatic mammals (cetaceans and sirenians) appear in its fossil record
only from the Miocene, and solely as freshwater forms (Antoine et al., 2017).
Meanwhile, during the Eocene, there were marine basins deeply intruding into
the South American continent and reaching its inner regions (Louterbach et al.,
2014); therefore, marine mammals could have inhabited Amazonia during that time. Here, we
report the first record of an Eocene marine mammal, namely, a cetacean of the
family Basilosauridae, from the northwestern Amazon region and discuss its
taxonomic and morphological affinities, as well as its environment in the
context of the geographic distribution of basilosaurids during the Eocene.
Diaz-Berenguer
et al. The Hind Limbs of Sobrarbesiren cardieli (Eocene, Northeastern Spain) and New Insights into the
Locomotion Capabilities of the Quadrupedal Sirenians. Journal of Mammalian
Evolution.
Link:
https://link.springer.com/article/10.1007%2Fs10914-019-09482-9
Abstract:
In the transition from a terrestrial to an aquatic environment, sirenian marine
mammals reduced and lost their hind limbs and developed a horizontal caudal
fin, the main propulsive organ in extant sirenians. Quadrupedal forms are only
known from the Eocene and are represented by three different clades: the
amphibious “prorastomids,” the aquatic quadrupedal protosirenids, and Sobrarbesiren
cardieli, a four-legged sirenian from the middle Eocene of Spain,
considered the sister taxon of the fully aquatic Dugongidae. This ecological
shift from terrestrial to an aquatic environment was naturally associated with
adaptations, among others, of the skeleton. However, sirenian hind limb bones
have been poorly studied because of the scarce material available in the fossil
record. Here, we describe in detail the hind limb bones of Sobrarbesiren,
analyzing their functional morphology and comparing them with other basal
sirenians and cetaceans, and with related terrestrial mammals such as proboscideans
and hyracoids. The hind limbs of Sobrarbesiren were capable of
a great variety of movements. Based on the presence of a strong sacroiliac
articulation, we propose that it swam by dorsoventral pelvic undulation
combined with pelvic paddling analogous to extant otters and the
“prorastomid” Pezosiren. We also conduct the first microanatomical
analysis of hind limb bones of an Eocene sirenian. Data reveal extreme inner
compactness in the Sobrarbesiren innominate and femur, with
the first description of osteosclerosis in an amniote innominate combined with
the highest degree of osteosclerosis observed in amniote femora. The results
confirm that the microanatomical changes precede the external morphological
changes in such ecological transitions. The process of adaptation of sirenians
to an aquatic life was thus a more complex process than previously thought.

Di
Celma et al. Allostratigraphy and paleontology of the lower Miocene Chilcatay
Formation in
the Zamaca area, East Pisco basin, southern Peru. Journal of Maps.
Link:
https://www.tandfonline.com/doi/full/10.1080/17445647.2019.1604439?af=R
Abstract:
Based on mapping of laterally traceable stratigraphic discontinuities, we
propose a high-resolution allostratigraphic scheme for one of the world’s
foremost fossil marine vertebrate Lagerstätten: the lower Miocene strata of the
Chilcatay Formation exposed along the
Ica River near Zamaca,
southern
Peru. Measured sections combined
with 1:10,000 scale mapping of a 24 km
2 area provide an
overview of the stratal architecture, as well as a general facies framework and
interpretation of the various depositional settings. As a whole, the Chilcatay
alloformation is bounded by the CE0.1 unconformity at the base and the PE0.0
unconformity at the top. An internal Chilcatay surface, termed CE0.2, splits
the alloformation into two distinct allomembers (Ct1 and Ct2). The Ct1
allomember comprises three facies associations recording deposition in
shoreface, offshore, and subaqueous delta settings. The Ct2 allomember
comprises two facies associations, recording deposition in shoreface and
offshore settings. Using these data, we place the rich marine vertebrate
assemblage in a precise spatial and stratigraphic framework. The
well-diversified vertebrate assemblage is dominated by cetaceans (mostly
odontocetes) and sharks (mostly lamniforms and carcharhiniforms); rays, bony
fish, and turtles are also present. Taxonomic novelties include the first
records of baleen whales, platanistids, and eurhinodelphinids from the Chilcatay
Formation.

Dominici et al. The awkward record of fossil whales. Earth Science Reviews.
Link: https://www.sciencedirect.com/science/article/abs/pii/S0012825219304301
Abstract: The habitat, actuopaleontology and global fossil record of crown
cetaceans, today occupying apex positions in the marine ecosystem, is
reviewed. A large Neogene- Quaternary dataset is built, covering the
time span of the evolutionary radiation of crown Odontoceti and
Mysticeti and including updated information on whale taxonomy,
chronostratigraphy, geography, paleoenvironment, taphonomy and size. We
outline an uneven chronostratigraphic and geographic distribution of
fossils, which influences our understanding of global diversity trends.
Notwithstanding the vast majority of whale carcasses sinks to bathyal
depths, the fossil record is mainly associated with shelf
paleoenvironments. The evolution of gigantic whales triggered the
radiation of whale-fall communities, including a global bone-eating
fauna that hampers the preservation of carcasses at bathyal depths. This
“Osedax” effect may explain the unexpected distribution of the fossil
record, particularly in the Pleistocene, when baleen whales became
gigantic and the ecosystem engineers they are today. A review of the
relative thickness of Pleistocene marine strata rules out artefacts of
the rock record. The distribution of taphonomic grades suggests that the
average skeletal completeness decreases during the Neogene and
Quaternary, consistently with an increased efficiency of bioeroders. The
frequency of complete and articulated skeletons is time-independent,
suggesting a control by sedimentation rates. Quality of the record is
expected to improve particularly from taxonomic studies dedicated to the
rich, but unexplored South american record, documenting the record of
little known productive regions, such as Africa, the Arctic and
Antarctica, and the taphonomy and stratigraphic paleobiology of old and
new findings.

Domning
and Beatty. Fossil Sirenia of the West Atlantic and Caribbean region. XII. Stegosiren
macei, gen. et sp. nov. Journal of Vertebrate Paleontology.
Link:
https://www.tandfonline.com/doi/abs/10.1080/02724634.2019.1650369
Abstract: Stegosiren
macei, a new genus and species of halitheriine dugongid from the
mid-Oligocene of South Carolina, U.S.A. (Ashley and Chandler Bridge formations,
late Rupelian–late Chattian), represents a stage of halitheriine evolution more
derived than that of the Old World early Oligocene Eosiren imenti and Halitherium
schinzii, but slightly less derived than the West Atlantic late
Oligocene Metaxytherium albifontanum. It is more comparable in
stage of evolution to its early Oligocene contemporaries Caribosiren
turneri and Priscosiren atlantica and may be a sister
taxon of these two. It is distinguished autapomorphically from all other
sirenians by a notably broadened frontal roof and a thickened anterior tip of
the frontal, which formed a butt joint with the premaxilla. Analogous
(independently evolved) joints in several other sirenians (principally
dugongines) are correlated with enlarged upper tusks thought to be used for
excavating seagrass rhizomes. This suggests that large tusks also may have been
present (although not preserved) in Stegosiren, which is only the
second halitheriine in which such a feature has been observed. Stegosiren
macei brings to at least seven the number of potentially sympatric
sirenian species lineages known from the West Atlantic-Caribbean Oligocene (six
or more from South Carolina alone). This extraordinary
sirenian diversity, unmatched elsewhere in the world, poses problems for
ecomorphology and feeding-niche partitioning.
Comments:
This marks the fourth named holotype specimen from the collections at our
museumm, the Mace Brown Museum of Natural History. This is yet another goddamn
sirenian from the Oligocene deposits here - we're already full - Priscosiren,
Metaxytherium albifontanum, Crenatosiren, Dioplotherium manigaulti - all of
these are known from the Oligocene here! The world's most diverse sea cow
assemblage.
Freschi
et al. Chronostratigraphic distribution of cetaceans in the Pliocene of
Northern
Apennines (Italy): palaeoecological
implications. Palaeogeography, Palaeoclimatology, Palaeoecology.
Link:
https://www.sciencedirect.com/science/article/abs/pii/S0031018218307156
Abstract:
A large number of cetacean fossils have been recovered through the time from
the Pliocene deposits in Northern Apennines thrust belt, on both the
Padan-Adriatic and the Tyrrhenian side. In this work, the cetaceans of
Castell'Arquato Basin (Padan-Adriatic side) have been placed within a
high-resolution chronostratigraphic framework available for this area. This
results in a better assessment of their evolutionary history and of the
palaeoecologic factors controlling their diversity and abundance through the
time. Our results document a greater diversity between 3.1 and 2.7 Ma time
interval, likely due to the development of eutrophic conditions during precessionally-driven
insolation maxima at 400 ka eccentricity maxima, which are recorded in deeper
depositional settings by sapropel clusters. Eutrophic conditions kept a complex
trophic chain, as currently observed in the Ligurian Sea (Corsican-Ligurian-Provençal Basin), an area characterized also by
upwelling. A reduction in both number and ecological diversity of cetacean taxa
is observed along the Pliocene-Pleistocene transition; this trend, related to
the onset of the Quaternary icehouse conditions, appears in good agreement with
the recently recognized extinction event that affecting the marine megafauna at
the end of the Pliocene.
Comments:
The merger of vertebrate paleontology and stratigraphy at its finest. This is
one of my favorite papers from the past few years. Just... a model paper for
discussing fossils, environments, and time.
Gaetan
et al. Prosqualodon australis (Cetacea: Odontoceti) from the
Early Miocene of Patagonia, Argentina: Redescription and Phylogenetic Analysis.
Ameghiniana.
Link:https://bioone.org/journals/ameghiniana/volume-56/issue-1/AMGH.21.11.2018.3208/Prosqualodon-australis-Cetacea--Odontoceti-from-the-Early-Miocene-of/10.5710/AMGH.21.11.2018.3208.short
Abstract:
We re-describe the holotype and referred specimen of Prosqualodon
australis, from the early Miocene of Gaiman Formation (Chubut Province, Argentina), and analyse its phylogenetic
position. Our study shows that this taxon is presently represented by an adult
and a juvenile individuals and we provide the first diagnosis of the species
based on the morphology of the anterior projection of the antorbital process
along with plesiomorphic dental features (such as well-marked heterodont teeth,
with posterior double rooted teeth in the maxilla, large, lateromedially compressed
teeth with long crowns and denticulated enamel). The total body length of P.
australis is estimated to be 2 m in the juvenile and 3 m in the adult
specimen. We included for the first time this species in a phylogenetic
analysis as an operational taxonomic unit itself, recovering P.
australis as a stem-odontocete and Prosqualodon as a
paraphyletic genus.
Comment:
A much needed re-description of Prosqualodon with new specimens. Sadly, no
earbones are known. The original fossils of the other species, Prosqualodon
davidis from Tasmania, have been tragically
lost.
Gibson
et al. Tupelocetus palmeri, a new species of protocetid whale (Mammalia,
Cetacea)
from the middle Eocene of South Carolina. Journal of Vertebrate Paleontology.
Link:
https://www.tandfonline.com/doi/full/10.1080/02724634.2018.1555165
Abstract:
We describe Tupelocetus palmeri, a new genus and species of
archaeocete whale, based on a single specimen from the Cross Member of the
middle Eocene Tupelo Bay Formation (Bartonian). The holotype consists of a
partial cranium with complete petrosals, posterior processes of tympanics, the
posterior ends of the nasals, a part of the right orbit, and what are
interpreted as the right and left P2s. The incomplete skull was collected from the
Martin Marietta Cross Quarry (Berkeley County, South Carolina), which has also furnished the
holotype of Carolinacetus gingerichi. The new species differs from
all other protocetids in having a deep cavity on the occiput, as well as having
the following combination of features: large nasal processes of frontals,
single-cusped P2, and premaxilla terminates at level of P2 or P3. A
phylogenetic analysis places T. palmeri in a large polytomy
along the cetacean stem, more closely related to crown Cetacea than Maiacetus, Artiocetus,
and Rodhocetus, but more basal than Georgiacetus, Babiacetus,
and Eocetus. Another member of this polytomy is Natchitochia
jonesi, and although specimens of Natchitochia and T.
palmeri do not share any elements, what is preserved suggests that
they are similar in size and among the largest of all protocetids. Although we
are unable to differentiate these two species, we outline future data that
could resolve this question.
Comment:
Not the most complete protocetid ever, but these poorly known whales are rare
as hen's teeth outside the Middle East - and this one has some of
the best preserved and prepared earbones of any protocetid. Also, from right
here in the Charleston area!
Gingerich
et al. Aegicetus gehennae, a new late Eocene protocetid (Cetacea, Archaeoceti)
from Wadi
Al Hitan, Egypt, and the transition to tailpowered swimming in whales. PLoS
One.
Link:
https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0225391
Abstract: Aegicetus
gehennae is a new African protocetid whale based on a partial skull
with much of an associated postcranial skeleton. The type specimen, Egyptian Geological Museum, Cairo [CGM] 60584, was found near the base
of the early-Priabonian-age (earliest late Eocene) Gehannam Formation of the
Wadi Al Hitan World Heritage Site in Egypt. The cranium is distinctive in
having ventrally-deflected exoccipitals. The vertebral column is complete from
cervical C1 through caudal Ca9, with a vertebral formula of 7:15:4:4:9+,
representing, respectively, the number of cervical, thoracic, lumbar, sacral,
and caudal vertebrae. CGM 60584 has two more
rib-bearing thoracic vertebrae than other known protocetids, and two fewer
lumbars. Sacral centra are unfused, and there is no defined auricular surface
on the ilium. Thus there was no weight-bearing sacroiliac joint. The sternum is
distinctive in being exceptionally broad and flat. The body weight of CGM 60584, a putative male, is
estimated to have been about 890 kg in life. Long bones of the fore and hind limbs
are shorter than expected for a protocetid of this size. Bones of the manus are
similar in length and more robust compared to those of the pes. A log vertebral
length profile for CGM 60584 parallels that of
middle Eocene Maiacetus inuus through the anterior and middle
thorax, but more posterior vertebrae are proportionally longer. Vertebral
elongation, loss of a sacroiliac articulation, and hind limb reduction indicate
that Aegicetus gehennae was more fully aquatic and less
specialized as a foot-powered swimmer than earlier protocetids. It is doubtful
that A. gehennae had a tail fluke, and the caudal
flattening known for basilosaurids is shorter relative to vertebral column
length than flattening associated with a fluke in any modern whale. Late
protocetids and basilosaurids had relatively long skeletons, longer than those
known earlier and later, and the middle-to-late Eocene transition from
foot-powered to tail-powered swimming seemingly involved some form of
mid-body-and-tail undulation.
Comment:
This new protocetid is a fairly derived one, and a very late one at that: it is
Priabonian, and overlaps in age with many basilosaurids. We don't know much
about the transition from four-legged protocetids to fully marine
basilosaurids, and ironically this whale may not tell us a whole lot since it
post-dates many basilosaurids anyway. That being said, it's got slightly
smaller hindlimbs than other protocetids
Govender.
Fossil cetaceans from Duinefontein (Koeberg) an early Pliocene site on the
southwestern Cape, South Africa. Palaeontologia Electronica.
Link: https://palaeo-electronica.org/content/2019/2398-koeberg-early-pliocene-cetacea
Abstract:
Koeberg is a Zanclean, subsurface locality on the west coast of South Africa, which was unearthed during the
excavations of the Koeberg Power Station foundations. Mysticetes and
odontocetes have been identified from this locality ca. 12 m
below the surface. The preservational nature prevents species level
identification in some instances. The balaenopterids are represented by
specimens assigned to ‘Balaenoptera’ borealina, Diunatans sp.
1 and Fragilicetus sp. while other specimens are only
identified as balaenopterids. The odontocetes have been referred to Odontoceti
indet. (possibly delphinids), a beaked whale (cf. Izikoziphius) and
a single phocoenid, cf.Australithax. There is undescribed odontocetes
most likely physeterids. The balaenopterids and odontocetes suggest a
connection of the west coast of South Africa to north Atlantic of Europe and
North
America,
and eastern South Pacific.

Kapur
et al. A Middle Miocene (~14 Ma) vertebrate assemblage from Palasava, Rapar
Taluka, Kutch (Kachchh) District, Gujarat State, western India. Historical Biology.
Link: https://www.tandfonline.com/doi/full/10.1080/08912963.2019.1648451
Abstract:
The present article aims to record a recently discovered middle Miocene
vertebrate assemblage (including marine and terrestrial mammals) from Palasava,
Kutch (Kachchh) District, Gujarat State, western India. The faunal assemblage comprising
fishes, chelonians, crocodiles, snakes, birds and mammals is significant in
terms of palaeoecology, palaeoenvironment and biostratigraphy of the region and
yields evidence concerning its biogeographic affinities linked to Neogene
Eustatic history. Palasava fauna together with previously known Neogene
(particularly Miocene) vertebrate assemblages from the Kutch region indicates the presence
of a riverine-estuarine system(s) linked to the sea, i.e. the depositional
centres were quite close to the sea. Furthermore, the Palasava vertebrate
assemblage (recorded herein) points towards a prevalence of warm, humid/wet,
tropical to sub-tropical environmental conditions. The terrestrial mammalian
assemblage from Palasava represents at least seven families with six of the
terrestrial mammalian taxa identified herein to the genus level, namely: Sanitherium,
Sivameryx, Brachypotherium, Zygolophodon, Gomphotherium, and Deinotherium.
The overall assemblage (including Zygolophodon and Deinotherium)
is indicative of a Middle Miocene, 14 ± 2 Ma (Langhian to
Serravallian) age for the sedimentary succession at Palasava. Furthermore, a
~ 14 Ma age for the Palasava fauna indicates a correlation to
regression cycle R5 in the Tagus Valley succession of Portugal.

Kienle
and Berta. The evolution of feeding strategies in phocid seals (Pinnipedia,
Phocidae).
Link:
https://www.tandfonline.com/doi/full/10.1080/02724634.2018.1559172
Abstract:
Adaptations for feeding underwater were crucial to the success of pinnipeds
(seals, sea lions, and walruses) in their transition from terrestrial to
aquatic habitats. Extant phocids (true seals) use multiple feeding
strategies—biting, filter, and suction feeding—to capture and consume prey, and
each strategy is associated with cranial, mandibular, and dental adaptations.
However, little is known about feeding strategies in stem pinnipeds. The
objectives of this study were to investigate feeding strategies used by some
extinct pinnipeds based on cranial and mandibular morphologies and use this framework
to examine the evolution of phocid feeding strategies. Three-dimensional
cranial and mandibular landmark data were collected from 249 extant and fossil
pinnipeds. Principal component analysis and canonical variate analysis were
performed to describe the major axes of variation and compare overlap of fossil
and extant taxa in morphospace. Stem pinnipeds had morphologies associated with
biting and filter feeding. Several fossil taxa were most similar to extant
biters, suggesting that biting was a common and important feeding strategy for
early phocids. One fossil taxon, Homiphoca capensis, was
potentially a filter feeder, because it consistently overlapped with extant
filter feeders in cranial morphospace. No fossil taxa had morphological
adaptations for suction feeding, indicating that suction feeding is a more
derived strategy in phocids. Extant phocids and their ancestors have cranial
and mandibular adaptations for multiple feeding strategies, which allowed these
animals to move into diverse aquatic niches and likely contributed to their
successful transition from terrestrial to aquatic ecosystems.

Kimura.
First squalodelphinid from the early Miocene of the Pacific realm in the
Northern Hemisphere. Journal of Vertebrate Paleontology.
Link: https://www.tandfonline.com/doi/full/10.1080/02724634.2018.1493595
Abstract:
Squalodelphinidae are moderately long-snouted dolphins from the late Oligocene
to middle Miocene epochs (Marx et al., 2016). This family includes only six
species in six genera. Recent studies have confirmed that Squalodelphinidae
belong to the superfamily Platanistoidea (e.g., Lambert et al., 2014, 2017;
Tanaka and Fordyce, 2015, 2016; Kimura and Barnes, 2016; Boersma et al., 2017;
Tanaka et al., 2017; Bianucci et al., 2018). The only surviving Platanistoidea
species, the South Asian river dolphin (Platanista gangetica), is distributed
in the Ganges, Brahmaputra-Megna, and
Karnaphuli-Sangu river systems of South Asia (Jefferson et al., 2008).
Several recent studies have described fossil platanistoids from Japan (e.g., Kimura et al., 2009, and
references therein; Kimura et al., 2013; Kimura and Barnes, 2016), which shed
light on the group’s paleobiogeography and evolution in this area. However, the
Platanistoidea fossil record from the wider Asian region is limited, and their
evolutionary history remains unclear. This is especially true for
Squalodelphinidae
because
their fossil record is relatively rare and diagnostic material of
squalodelphinids has been exclusively reported from the Atlantic and Pacific coasts of South America, the eastern coast
of North America, and Europe (Tanaka and Fordyce, 2014;
Bianucci et al., 2015, 2018). Here, I report the first squalodelphinid from the
Pacific region of the Northern Hemisphere: beautifully
preserved
ear bones recovered from the lower Miocene Iwamura Group, Gifu Prefecture, Japan. This new fossil expands our
knowledge of the paleobiogeography and evolution of the enigmatic odontocete
family Squalodelphinidae.
Comments:
See also the paper by Margot Nelson et al., reporting a partial squalodelphinid
skull from the Clallam Formation of Washington
USA: https://bioone.org/journals/Journal-of-Vertebrate-Paleontology/volume-38/issue-2/02724634.2017.1428197/First-Occurrence-of-a-Squalodelphinid-Cetacea-Odontoceti-from-the-Early/10.1080/02724634.2017.1428197.short
Kimura
and Hasegawa. New specimen of Joumocetus shimizui from the Miocene Haraichi
Formation, Annaka Group, Gunma Prefecture, Japan. Bulletin of the Gunma Museum of Natural History.
Link: http://www.gmnh.pref.gunma.jp/research_no23
Abstract:
A new mysticete fossil was recovered from the Haraichi Formation, Annaka Group,
Gunma
Prefecture, Japan. The specimen includes incomplete cranium with
periotics and tympanic bulla. The specimen represents the following
morphological characters and here we describe the specimen as Joumocetus
shimizui: parietals are widely exposed at vertex, supraoccipital shield is
triangular in outline with a pointed apex, anterior process of the periotic is
short and compressed transversely, and, although it is unclear due to brakeage,
nasal is possibly located in part anterior to the level of the preorbital angle
of the supraorbital process of the frontal. The horizon from which the specimen
was found is stratigraphically between the Kitamura and Baba tuffs. In previous
study, the geological ages of the Kitamura and the Baba tuffs was dated as
11.79 ± 0.08 Ma (biotite) and 11.26 ± 0.09 Ma (biotite)/11.29 ± 0.12 Ma
(sanidine) respectively. This new specimen elucidates the morphology of J.
shimizui which was not preserved in the holotype specimen of the species.
Comments:
In Japanese, but well-illustrated as is always the case with Kimura's work, and
adds some new character codings to the early cetotheriid Joumocetus.
Kimura
and Hasegawa. A new species of Kentriodon (Cetacea,
Odontoceti, Kentriodontidae) from the Miocene of Japan.
Link: https://www.tandfonline.com/doi/full/10.1080/02724634.2019.1566739
Abstract:
Kentriodontids are small- to medium-sized odontocetes with a wide geographic
range that flourished during the Miocene. They are closely related to crown
Delphinida. Seven fossil kentriodontid specimens have been recovered from the
Haraichi Formation, Annaka Group (latest middle/earliest late Miocene,
Serravallian/Tortonian), Gunma Prefecture, Japan. We describe and diagnose a new
species of the genus Kentriodon. Kentriodon nakajimai,
sp. nov., has a larger fossa for the hamular and preorbital lobes of the
pterygoid sinuses, which suggests that the species was more highly adapted for
diving than other Kentriodon species. Asymmetric development
of left and right maxillary crests indicates asymmetric development of the
facial muscles attached to the melon. If this is the case, it suggests
that K. nakajimai possessed a more sophisticated echolocation
system than other kentriodontids. Phylogenetic analysis suggests that K.
nakajimai is most closely related to K. obscurus and
the genus Kentriodon may have originated in the Pacific and
then dispersed into the Atlantic several times.
Comments:
What a spectacular collection of fossils - quite a sample size! The second
species of Kentriodon reported from Japan, and very likely the best known
species to date.
Lambert
and Bianucci. How to break a sperm whale’s teeth: dental damage in a large
Miocene physeteroid from the North Sea basin. Journal of Vertebrate
Paleontology.
Link:
https://www.tandfonline.com/doi/full/10.1080/02724634.2019.1660987
Abstract:
In contrast to the suction-feeding, predominantly teuthophagous extant sperm
whale, several Miocene physeteroids display proportionally larger teeth, deeply
embedded in both upper and lower jaws. Together with other osteological
features, these differences lead to the functional interpretation of these taxa
as macroraptorial predators, using their teeth to capture and process large
marine vertebrates. However, the assumption that strong forces applied to
macroraptorial physeteroid teeth during powerful bites and contacts with bone
material should result in major dental damage has not yet been tested. In the
present work, we analyzed a large collection of physeteroid teeth with an
enameled crown from the Miocene of the North Sea Basin. We especially focused on a set
of 45 teeth of Scaldicetus caretti discovered in Antwerp (Belgium, southern North Sea Basin) and tentatively dated to the
Tortonian (early late Miocene). Visual inspection and computed tomography (CT)
scans revealed dental damage, including wear and breaks. The latter could be
interpreted as chipping fractures, occurring along the crown, and vertical root
fractures, observed along the apical part of the massive root. Chipping
fractures are most likely due to contacts with hard material, whereas vertical
root fractures may result from the application of strong and repetitive bite
forces and/or contacts with hard material. Such results further support the
interpretation of a series of Miocene physeteroids with proportionally large
teeth as macroraptorial (rather than suction-feeding) top predators.
Considering the size of the teeth of S. caretti, its most likely
prey items were other large marine vertebrates.
Comments:
Every paleocetologist is familiar with tooth breakage and feeding -
macropredatory whales bite down on a bony prey item like a turtle or a seal and
the tooth crown breaks. There's a little bit of that in here, which is what I
was expecting. What I was not expecting was breakage of the tooth roots during
biting!!!
Lambert et al. An Amphibious Whale from the Middle Eocene of Peru Reveals Early South Pacific Dispersal of Quadrupedal Cetaceans. Current Biology.
Link: https://www.cell.com/current-biology/fulltext/S0960-9822(19)30220-9?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0960982219302209%3Fshowall%3Dtrue
Abstract: Cetaceans originated in south Asia more than 50 million years ago (mya), from a small quadrupedal artiodactyl ancestor. Amphibious whales gradually dispersed westward along North Africa and arrived in North America before 41.2 mya. However, fossil evidence on when, through which pathway, and under which locomotion abilities these early whales reached the New World is fragmentary and contentious. Peregocetus pacificus gen. et sp. nov. is a new protocetid cetacean discovered in middle Eocene (42.6 mya) marine deposits of coastal Peru, which constitutes the first indisputable quadrupedal whale record from the Pacific Ocean and the Southern Hemisphere. Preserving the mandibles and most of the postcranial skeleton, this unique four-limbed whale bore caudal vertebrae with bifurcated and anteroposteriorly expanded transverse processes, like those of beavers and otters, suggesting a significant contribution of the tail during swimming. The fore- and hind-limb proportions roughly similar to geologically older quadrupedal whales from India and Pakistan, the pelvis being firmly attached to the sacrum, an insertion fossa for the round ligament on the femur, and the retention of small hooves with a flat anteroventral tip at fingers and toes indicate that Peregocetus was still capable of standing and even walking on land. This new record from the southeastern Pacific demonstrates that early quadrupedal whales crossed the South Atlantic and nearly attained a circum-equatorial distribution with a combination of terrestrial and aquatic locomotion abilities less than 10 million years after their origin and probably before a northward
dispersal toward higher North American latitudes.
Comments: Not only is this new protocetid the first named protocetid from the Pacific, but it's also got pretty great limb material!
Lambert et al. Yaquinacetus meadi, a new latest Oligocene–early Miocene dolphin (Cetacea, Odontoceti, Squaloziphiidae, fam. nov.) from the Nye Mudstone (Oregon, U.S.A.). Journal of Vertebrate Paleontology.
Link: https://www.tandfonline.com/doi/full/10.1080/02724634.2018.1559174
Abstract: Represented by a nearly complete cranium with associated mandible, teeth, and vertebrae, Yaquinacetus meadi is a new genus and species of archaic homodont odontocete from the latest Oligocene–early Miocene (24–19.2 Ma) of Oregon, U.S.A. The new species is characterized by a moderately elongated rostrum bearing approximately 51 alveoli per tooth row and a knob-like, rectangular vertex. Together with Squaloziphius emlongi from the early Miocene of Washington State, Y. meadi constitutes a new odontocete family, Squaloziphiidae, fam. nov., diagnosed by a unique combination of characters, including transversely wide dorsal opening of the mesorostral groove at base of rostrum, followed posteriorly by an abrupt narrowing; thickened lateral margin of the maxilla in the antorbital region making a long and laterally concave crest; and massive, anteroposteriorly and ventrally long postglenoid process of the squamosal. Although sharing with Ziphiidae the presence of transverse premaxillary crests on the vertex, Squaloziphiidae differs in the pterygoid sinus fossa being shorter anteriorly and ventrally; the tubercule of the malleus being less reduced; and lacking a pair of enlarged alveoli for mandibular tusks. Our phylogenetic analysis confirms the sister-group relationship between S. emlongi and Y. meadi, either as late diverging stem odontocetes or as early crown odontocetes, but distant from Ziphiidae. These results confirm the northeastern Pacific as a center of diversification for several groups of archaic homodont odontocetes during the late Oligocene–early Miocene.
Comments: This is only the second species of squaloziphiid named since Squaloziphius was first named in 1991. it's also got the first earbones for the family (and, the clade Squaloziphiidae is named). Based on a beautiful skull collected by Doug Emlong from the Nye Mudstone of Oregon.
Leslie et al. Norrisanima
miocaena, a new generic name and redescription of a stem
balaenopteroid mysticete (Mammalia, Cetacea) from the Miocene of California.
PeerJ.
Link:
https://peerj.com/articles/7629/
Abstract:
Rorqual whales are among the most species rich group of baleen whales
(or mysticetes) alive today, yet the monophyly of the traditional grouping
(i.e., Balaenopteridae) remains unclear. Additionally, many fossil mysticetes
putatively assigned to either Balaenopteridae or Balaenopteroidea may actually
belong to stem lineages, although many of these fossil taxa suffer from
inadequate descriptions of fragmentary skeletal material. Here we provide a
redescription of the holotype of
Megaptera miocaena, a fossil
balaenopteroid from the Monterey Formation of California, which consists of a
partial cranium, a fragment of the rostrum, a single vertebra, and both
tympanoperiotics.
Kellogg (1922) assigned the type specimen to the genus
Megaptera
Gray (1846), on the basis of its broad similarities to distinctive
traits in the cranium of extant humpback whales (
Megaptera novaeangliae
(
Borowski, 1781)). Subsequent phylogenetic analyses have found
these two species as sister taxa in morphological datasets alone; the most
recent systematic analyses using both molecular and morphological data sets
place
Megaptera miocaena as a stem balaenopteroid unrelated to humpback whales.
Here, we redescribe the type specimen of
Megaptera miocaena in the
context of other fossil balaenopteroids discovered nearly a century since
Kellogg’s original description and provide a morphological basis for
discriminating it from
Megaptera novaeangliae. We also provide a new
generic name and recombine the taxon as
Norrisanima miocaena, gen. nov.,
to reflect its phylogenetic position outside of crown Balaenopteroidea,
unrelated to extant
Megaptera. Lastly, we refine the stratigraphic age
of
Norrisanima miocaena, based on associated microfossils to a Tortonian
age (7.6–7.3 Ma), which carries implications for understanding the origin of
key features associated with feeding and body size evolution in this group of
whales.
Comments: A much needed redescription of
"Megaptera" miocaena, originally published by Kellogg in the 1920s.
Many more fossils of Norrisanima, perhaps not N. miocaena, are present in
collections at UCMP, SDNHM, and LACM, so
it's nice to finally apply a genus name other than "Megaptera". I
find the images of the skull - derived from 3D models - washed out and
difficult to see, however.
Loch et al. Enamel
Microstructure in Eocene Cetaceans from Antarctica (Archaeoceti and Mysticeti). Journal of Mammalian Evolution.
Link:
https://link.springer.com/article/10.1007/s10914-018-09456-3
Abstract:
Modern baleen whales have no adult teeth, whereas dolphins and porpoises
have a homodont and polydont dentition, with simplified enamel microstructure.
However, archaic cetaceans (archaeocetes and early mysticetes and odontocetes)
had a complex and ornamented dentition, with complex enamel microstructure as
in terrestrial mammals. This study describes the morphology of teeth and enamel
microstructure in two fossil cetaceans from Antarctica:
a basilosaurid archaeocete from the La Meseta Formation (middle Eocene); and Llanocetus
sp. from the Submeseta Formation (late Eocene), one of the oldest mysticetes
known. The two teeth analyzed were lower premolars, with transversely
compressed triangular crowns composed of a main cusp and accessory denticles.
The enamel microstructure of the basilosaurid and Llanocetus sp. is
prismatic with Hunter-Schreger bands (HSB)
and an outer zone of radial enamel. In the basilosaurid, the enamel is
relatively thin and measures 150–180 μm, whereas in Llanocetus sp.
it is considerably thicker, measuring 830–890 μm in the cusp area and
350–380 μm near the crown base. This is one of the thickest enamel layers
among cetaceans, extinct and living. Structures resembling enamel tufts and
lamellae were observed in both fossils at the enamel-dentine junction (EDJ) and
extending along the thickness of the enamel layer, respectively. The presence
of HSB and biomechanical reinforcing
structures such as tufts and lamellae suggests prominent occlusal loads during
feeding, consistent with raptorial feeding habits. Despite the simplification
or absence of teeth in modern cetaceans, their ancestors had complex posterior
teeth typical of most mammals, with a moderately thick enamel layer with prominent
HSB.
Comments: There is
a raging debate as to what toothed mysticetes ate, and this paper adds some
fuel to that fire. This paper reports enamel features of Llanocetus, which has
unusually thick enamel - suggestive of powerful bite force rather than
reduction of tooth function related to either suction feeding or filter
feeding.
Marx et al. Like phoenix from the ashes: How modern baleen whales arose from a fossil “dark age”. Acta Palaeontologica Polonica.
Link: https://www.app.pan.pl/article/item/app005752018.html
Abstract: The evolution of baleen whales (Mysticeti), the largest animals on Earth, was punctuated by a pivotal turnover event. Following their emergence around 36 million years (Ma), mysticetes diversified into a disparate range of toothed and toothless species until 23 Ma, but then nearly vanished from the global fossil record for the next five million years. Following this early Miocene “dark age”, toothless mysticetes spectacularly reappeared around 18–17 Ma, whereas toothed mysticetes had gone entirely extinct. Here, we suggest that this turnover event reflects a change in mysticete habitat occupancy. Using the well-sampled record of Australasia as a case study, we show that Oligocene pre-“dark age” mysticetes formed distinct coastal and offshore assemblages, dominated by small (2–4 m), ecologically disparate toothed species, and larger (5–6 m) toothless filter feeders, respectively. Environmental change around the Oligocene–Miocene boundary led to the decline of the endemic coastal assemblages, leaving nearshore deposits virtually devoid of mysticetes. Filter feeders persisted offshore and subsequently re-invaded coastal habitats during the mid-Miocene Climatic Optimum, thus establishing the modern, cosmopolitan mysticete fauna.
Comments: One of Felix's corniest paper titles by far (we love you Felix). A quirky paper on a very poorly understood hitherto underappreciated aspect of the baleen whale fossil record: it sort of ceases to exist for about a 3-4 million year period (Aquitanian stage) yet dolphins are quite common in this interval, so it can't readily be explained away by time/rock bias. Felix started talking my ear off about this problem when we were Ph.D. students at Otago, and I'm glad to see it published. Don't forget the Supp Info though: they report the first eomysticetids from Australia!
Marx et al. Gigantic mysticete predators roamed the Eocene Southern Ocean. Antarctic Science.
Link: https://www.cambridge.org/core/journals/antarctic-science/article/gigantic-mysticete-predators-roamed-the-eocene-southern-ocean/0EEFC32753A8909BC4E7C134F5AEA6AE
Abstract: Modern baleen whales (Mysticeti), the largest animals on Earth, arose from small ancestors around 36.4 million years ago (Ma). True gigantism is thought to have arisen late in mysticete history, with species exceeding 10 m unknown prior to 8 Ma. This view is challenged by new fossils from
Seymour Island (Isla Marambio),
Antarctica, which suggest that enormous whales once roamed the Southern Ocean during the Late Eocene (
c. 34 Ma). The new material hints at an unknown species of the archaic mysticete
Llanocetus with a total body length of up to 12 m. The latter is comparable to that of extant Omura's whales (
Balaenoptera omurai Wada
et al. 2003), and suggests that gigantism has been a re-occurring feature of mysticetes since their very origin. Functional analysis including sharpness and dental wear implies an at least partly raptorial feeding strategy, starkly contrasting with the filtering habit of living whales. The new material markedly expands the size range of archaic mysticetes, and demonstrates that whales achieved considerable disparity shortly after their origin.
Comments: Tantalizing evidence in the form of huge teeth of Llanocetus, suggesting an even larger species than Llanocetus denticrenatus ranging up to 12 meters in length. However, since these are just isolated teeth, it's admittedly possible that this was a much larger-toothed species of Llanocetus. We don't have a skull for this taxon, so it's difficult to really say much about its size or feeding ecology.
Marx et al. A large Late Miocene cetotheriid (Cetacea, Mysticeti) from the Netherlands clarifies the status of Tranatocetidae.
Link: https://peerj.com/articles/6426/
Abstract: Cetotheriidae are a group of small baleen whales (Mysticeti) that evolved alongside modern rorquals. They once enjoyed a nearly global distribution, but then largely went extinct during the Plio-Pleistocene. After languishing as a wastebasket taxon for more than a century, the concept of Cetotheriidae is now well established. Nevertheless, the clade remains notable for its variability, and its scope remains in flux. In particular, the recent referral of several traditional cetotheriids to a new and seemingly unrelated family, Tranatocetidae, has created major phylogenetic uncertainty. Here, we describe a new species of
Tranatocetus, the type of Tranatocetidae, from the Late Miocene of the
Netherlands.
Tranatocetus maregermanicum sp. nov. clarifies several of the traits previously ascribed to this genus, and reveals distinctive auditory and mandibular morphologies suggesting cetotheriid affinities. This interpretation is supported by a large phylogenetic analysis, which mingles cetotheriids and tranatocetids within a unified clade. As a result, we suggest that both groups should be reintegrated into the single family Cetotheriidae.
Comments: Another welcome discovery from the North Sea - Trantocetidae, erected only a few years ago, is now found to be synonymous with the Cetotheriidae - something I suspected for a while. this study has one of the best resolved phylogenies of Mysticeti yet (though I'm not sure about Caperea).
Matsui and Tsuihiji. The phylogeny of
desmostylians revisited: proposal of new clades based on robust phylogenetic
hypotheses.
Link:
https://peerj.com/articles/7430/
Abstract: Desmostylia
is a clade of extinct aquatic mammals with no living members. Today, this clade
is considered belonging to either Afrotheria or Perissodactyla. In the
currently-accepted taxonomic scheme, Desmostylia includes two families, 10 to
12 genera, and 13–14 species. There have been relatively few phylogenetic
analyses published on desmostylian interrelationship compared to other
vertebrate taxa, and two main, alternative phylogenetic hypotheses have been
proposed in previous studies. One major problem with those previous studies is
that the numbers of characters and OTUs were small. In this study, we analyzed
the phylogenetic interrelationship of Desmostylia based on a new data matrix
that includes larger numbers of characters and taxa than in any previous
studies. The new data matrix was compiled mainly based on data matrices of
previous studies and included three outgroups and 13 desmostylian ingroup taxa.
Analyses were carried out using five kinds of parsimonious methods. Strict
consensus trees of the most parsimonious topologies obtained in all analyses
supported the monophyly of Desmostylidae and paraphyly of traditional
Paleoparadoxiidae. Based on these results, we propose phylogenetic definitions
of the clades Desmostylidae and Paleoparadoxiidae based on common ancestry.
Comments: Perhaps
the best phylogenetic analysis of desmos to date. Sadly, my favorite genus name
for the clade, Vanderhoofius is no more: now recombined as Desmostylus
coalingensis. Fortunately, Cornwallius, my second favorite genus name is still
accepted.
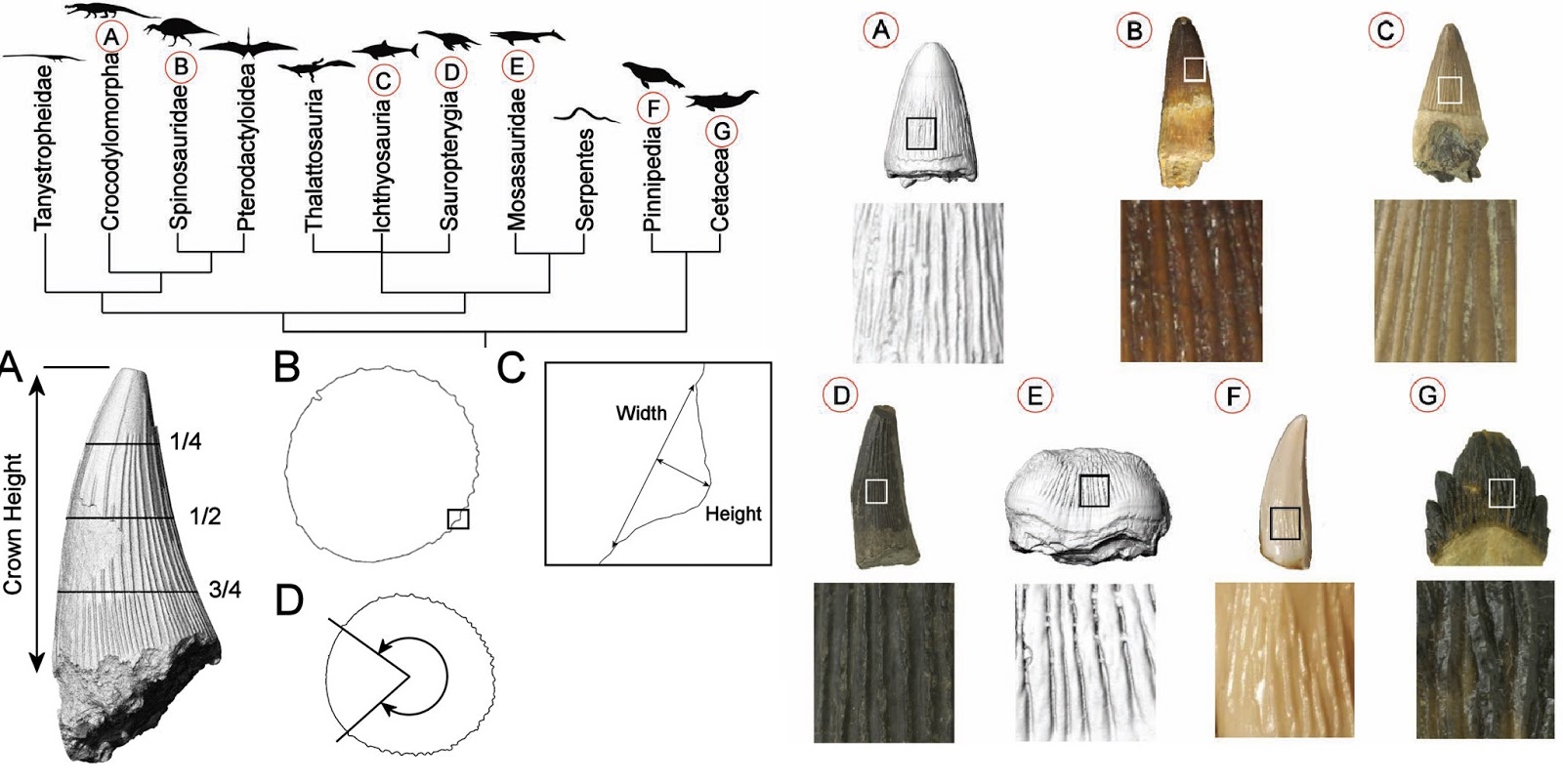
McCurry
et al. The repeated evolution of dental apicobasal ridges inaquatic-feeding
mammals and reptiles.
Link:
https://academic.oup.com/biolinnean/article/127/2/245/5427318
Abstract:
Since the Permian, Earth’s aquatic ecosystems have been ecologically
dominated by numerous lineages of predatory amniotes. Many of these groups
evolved elevated ridges of enamel that run down the apical–basal axis of their
teeth, referred to here as apicobasal ridges. This trait is commonly used as a
taxonomic tool to identify fossil species and higher groupings, but the
function of the ridges and their associated ecological significance are poorly
understood. Here, we aim to clarify the phylogenetic distribution of apicobasal
ridges among amniotes and to examine how the morphology of apicobasal ridges
varies across species. We show that these ridges have evolved independently
numerous times and are almost exclusively found in aquatic-feeding species.
Ridge morphology varies, including tall, pronounced ridges, low, undulating
ridges and interweaving ridges. Their internal structure also varies from tooth
crowns with locally thickened enamel to undulating enamel–dentine interface. We
assess the relative merits of different hypothetical functions of the ridges
and propose that although apicobasal ridges might provide some strengthening of
the tooth, their morphology and pattern of evolution do not indicate that this
is their primary function. Instead, we suggest that apicobasal ridges serve to
increase the efficiency of puncture, grip and/or removal.
Comments:
I was not expecting tooth shape/texture to be apparently decoupled from
internal structure - though this study does not examine enamel ultrastructure,
which is a necessary test of the central hypothesis: that enamel
"fluting" as a shape is important for piercing the tissues of prey
animals and not, for example, distributing stress from point loading.
Regardless, an interesting and thought provoking study.
Muizon
et al. Mystacodon selenensis, the earliest known toothed
mysticete (Cetacea, Mammalia) from the late Eocene of Peru: anatomy, phylogeny,
and feeding adaptations. Geodiversitas.
Link:
https://bioone.org/journals/Geodiversitas/volume-41/issue-1/geodiversitas2019v41a11/Mystacodon-selenensis-the-earliest-known-toothed-mysticete-Cetacea-Mammalia-from/10.5252/geodiversitas2019v41a11.short
Abstract:
Mystacodon selenensis Lambert, Martínez-Cáceres, Bianucci, Di
Celma, Salas-Gismondi, Steurbaut, Urbina & Muizon, 2017 is a toothed
mysticete that represents the earliest member of the suborder in the current
state of knowledge. Its holotype is a relatively complete skeleton from the
upper Eocene (early Priabonian, c. 36.4 Ma) Yumaque Member of the
Paracas Formation from the southern coast of Peru.
The thorough description of this specimen is presented here and reveals
numerous similarities with the contemporaneous basilosaurids including the
retention of an innominate that originally articulated to the unpreserved hind
limb. However, several characters of M. selenensis clearly relate this
taxon to the mysticetes, such as the large palate with a dorsoventrally
flattened rostrum, the posterior extension of the palate with an infraorbital
plate of the maxilla, the shortening of the premaxillary part of the rostrum,
the zygomatic process of the squamosal being closely apposed to the postorbital
process of the frontal, and the humeral head being oriented more proximally
than posteriorly. A parsimony analysis retrieves Mystacodon as the
earliest diverging branch of the Mysticeti with no close phylogenetic
relationship with Llanocetus the second oldest known mysticete (c.
34.2 Ma). The dental formula of M. selenensis is that of basilosaurids
(I 3/3, C 1/1, P 4/4, M 2/3). The anterior teeth (incisors and canine) are
distinctly proportionally smaller than in basilosaurids, whereas the cheek
teeth are very close in relative length, but are relatively larger than in most
other toothed mysticetes (except Coronodon). The large cheek teeth of Mystacodon
suggest a raptorial feeding strategy, probably assisted with some degree of
suction, as indicated by the large size of the palate. The anterior teeth of
the holotype display a subhorizontal apical wear facet and the cheek teeth a
moderately sloping wear surface, differing from the subvertical attrition
facets of basilosaurids. This pattern suggests an efficient dental abrasion
resulting from feeding upon abrasive food items or/and from the ingestion of
sediment during prey capture, which could indicate some degree of bottom
feeding. On the forelimb, the size and orientation of the acromion, the great
length of the deltopectoral crest, the massiveness of the olecranon of the
ulna, and the strong radial anterior process indicate powerful shoulder
movements, which suggest an active use of the forelimb when foraging for food
on the sea floor. The robustness of digits and the pachyosteosclerosis of ribs
with pestle-like distal end corroborate such a scenario. Mystacodon
selenensis represents a first step in the evolutionary history of feeding
adaptations of early mysticetes; the latter are likely to have experimented an
abundant set of feeding strategies and were probably very eclectic in prey
choice and capture before hyperspecialized filter feeding became widespread in
the suborder.
Comments: Great followup to the initial publication of the
earliest toothed mysticete Mystacodon in 2017 - well illustrated, exhaustive
paleobiological investigation.
Orihuela et al. First cranial endocasts of early Miocene
sirenians (Dugongidae) from the West Indies.
Journal of Vertebrate Paleontology.
Link:
https://www.tandfonline.com/doi/full/10.1080/02724634.2019.1584565
Abstract: We report and describe the
first sirenian endocranial casts from the West Indies
based on three specimens collected from two quarries of the upper
Oligocene–lower Miocene Colón Formation, in the province
of Matanzas, western Cuba.
We assign them to Dugongidae incertae sedis, based on a phylogenetic analysis
of fossil and extant sirenians. Thus, these new specimens provide a unique
opportunity to describe the endocranial neuroanatomy of this family. The
endocasts are suggestive of dugongids with limited vision and olfaction, based
on the diminished olfactory and optic nerves. Additionally, we provide a
geological reinterpretation of the Colón Formation and its paleoecological
setting. Altogether, these data provide further insight into the diversity and
evolution of sirenians, especially Caribbean dugongs.
Paolucci et al. Diaphorocetus poucheti (Cetacea, Odontoceti, Physeteroidea) from Patagonia, Argentina: one of the earliest sperm whales. Journal of Systematic Palaeontology.
Link: https://www.tandfonline.com/doi/full/10.1080/14772019.2019.1605544
Abstract: Sperm whales (Physeteroidea) are the basal-most surviving lineage of odontocetes, represented today by just three highly specialized, deep-diving suction feeders. By contrast, extinct sperm whales were relatively diverse, reflecting a major Miocene diversification into various suction feeding and macroraptorial forms. The beginnings of this diversification, however, remain poorly understood. The Atlantic coast of South America provides a crucial window into early physeteroid evolution and has yielded some of the oldest species known from cranial material, Idiorophus patagonicus and Diaphorocetus poucheti – both of which are in need of re-description and phylogenetic reappraisal. Here, we re-examine Diaphorocetus in detail and, in light of its complex taxonomic history, declare it a nomen protectum. Phylogenetically, the species forms part of a polytomy including ‘Aulophyseter’ rionegresis and the two crown lineages (Physeteridae and Kogiidae) and demonstrates that facial asymmetry and a clearly defined supracranial basin have characterized this lineage for at least 20 Ma. With a total body length of 3.5–4 m, Diaphorocetus is one of the smallest physeteroids yet known. Its cranial morphology hints at an intermediate raptorial/suction feeding strategy and it has a moderately developed spermaceti organ and junk.
Comments: Hot on the heels of other papers redescribing critical early Miocene cetaceans from Patagonia - Phoberodon, Morenocetus, Prosqualodon - and now this early sperm whale.
Peri et al. An Inticetus-like (Cetacea: Odontoceti) postcanine tooth from the Pietra leccese (Miocene, southeastern Italy) and its palaeobiogeographical implications, Neues Jahrbuch fur Geologie und Palaontologie.
Link: https://www.ingentaconnect.com/content/schweiz/njbgeol/2019/00000291/00000002/art00007
Abstract:
We report on an isolated cetacean postcanine tooth that was collected
close to the village of Melpignano (Lecce Province, Apulia region) from the
Miocene "Pietra leccese" formation of southeastern Italy. This tooth
exhibits a transversely compressed and roughly semi-circular crown featuring
several large, broad-based accessory denticles that are arranged radially.
Dental enamel ornamentation is limited to faint subvertical grooves, and a
slight subvertical incision just below the base of the crown suggests that it
was double-rooted. Our comparisons allow us to identify the Melpignano specimen
as belonging to a heterodont dolphin close to Inticetus vertizi, the
only named member of the archaic odontocete family Inticetidae, which has been
recently described from Burdigalian (early Miocene) strata of the Chilcatay
Formation of southern Peru.
This find highlights the elusive presence of Inticetus-like toothed
whales in the Mediterranean region during the Miocene. We then review the
geographic distribution of fossil remains of Inticetus-like cetaceans,
which includes records from the Miocene of Peru, North
Carolina (eastern U.
S. A.), the Atlantic coast of France,
and south-eastern Italy.
We hypothesise that inticetids dispersed via the Central American Seaway, which
allowed faunal interchanges between the south-eastern Pacific and the northern
Atlantic/Mediterranean realms until latest Miocene times. In conclusion, the
finding of an Inticetus -like tooth in Miocene beds in Italy
suggests that our knowledge on the past distribution of inticetid cetaceans is
far from being exhaustive.
Comments: One of the only papers to cite my paper on
Phococetus teeth from North Carolina!
Sigh...
Peri et al. Reduction of olfactory and respiratory turbinates in the transition of whales from land to sea: the semiaquatic middle Eocene Aegyptocetus tarfa. Journal of Anatomy.
Link: https://onlinelibrary.wiley.com/doi/full/10.1111/joa.13088
Abstract: Ethmoturbinates, nasoturbinates,
and maxilloturbinates are well developed in the narial tract of land‐dwelling
artiodactyls ancestral to whales, but these are greatly reduced or lost
entirely in modern whales. Aegyptocetus tarfa is a semiaquatic
protocetid from the middle Eocene of Egypt. Computed axial tomography scans of
the skull show that A. tarfa retained all three sets of turbinates
like a land mammal. It is intermediate between terrestrial artiodactyls and
aquatic whales in reduction of the turbinates. Ethmoturbinates in A. tarfa
have 26% of the surface area expected for an artiodactyl. These have an
olfactory function and indicate that early whales retained a sense of smell in
the transition from land to sea. Maxilloturbinates in A. tarfa have
6% of the surface area expected for an artiodactyl. These have a respiratory
function and their markedly reduced size suggests that rapid inhalation and
exhalation was already more important than warming and humidifying air, in
contrast to extant land mammals. Finally, the maxilloturbinates of A. tarfa,
although greatly reduced, still show some degree of similarity to those of
artiodactyls, supporting the phylogenetic affinity of cetaceans and
artiodactyls based on morphological and molecular evidence.

Powell
et al. Geology and Paleontology of the Late Miocene Wilson Grove Formation at Bloomfield Quarry, Sonoma County, California. USGS Professional
Publications.
Link:
https://pubs.usgs.gov/sir/2019/5021/sir20195021.pdf
Abstract:
An extensive fauna of at least 77 taxa is reported from the basal Wilson
Grove Formation in a small quarry just north of the town of Bloomfield,
Sonoma County, California.
The fauna represents intertidal to shallow subtidal water depths and water
temperatures interpreted from the fauna, consistent with the latitude of the
fossil locality (37° north) during the late Miocene. The fauna from Bloomfield
Quarry is unusually large and diverse from such a small area. It consists of
thousands of specimens of 4 brachiopod, 42 mollusk (28 bivalves and 14
gastropods), 6 arthropod (1 crab, 1 shrimp, and 4 barnacles), and 25 vertebrate
(3 sharks, 1 ray, 8 bony fishes, 9 marine mammals, and 4 birds) taxa. Unusual
in the fauna is the abundant and diverse brachiopod fauna, the diverse barnacle
fauna, which was described previously, and the extensive and diverse vertebrate
fauna. Most significant among the vertebrates is the walrus fauna, which is the
most diverse assemblage of walrus yet reported worldwide from a single
locality. A single strontium (Sr) isotope age determination of about 8 million
years (megaannum, Ma) from a pectinid mollusk is consistent with a new age
determination of the overlying, informally named Roblar tuff as described by
SarnaWojcicki in 1992 (6.203±0.011 Ma) and previously reported age
determinations (recalculated here) from basalt (9.27±0.06 Ma) underlying these
deposits. The Roblar tuff at Bloomfield Quarry can be correlated with other
sites, including the Delgada Fan offshore northern California
and the Coalinga anticline in California’s
Central Valley. These age determinations conform with
the “Jacalitos” California
provincial molluscan stage age, the Hemphillian North American Land Mammal age
determined from the fossils, and is part of the International Tortonian Stage
of the Miocene.
Comments:
This paper was a long time coming - I wrote the descriptions for this the month
before I got married to Sarah back in 2011. Nevertheless it boasts the most
diverse walrus fossil assemblage in the world: five taxa! Imagotaria,
Dusignathus, Gomphotaria, Pontolis, and an odobenine. This is also one of the
most exhaustive reports on a fossil site from the west coast: stratigraphy,
geochronology, invertebrate and vertebrate paleontology, you name it.
Everything we know about this locality.
Racicot et al. Evidence
for convergent evolution of ultrasonic hearing in toothed whales (Cetacea: Odontoceti).
Biology Letters.
Link:
https://royalsocietypublishing.org/doi/full/10.1098/rsbl.2019.0083
Abstract:
Toothed whales (Cetacea: Odontoceti) are the most diverse group of
modern cetaceans, originating during the Eocene/Oligocene transition approximately
38 Ma. All extant odontocetes echolocate; a single origin for this behaviour is
supported by a unique facial source for ultrasonic vocalizations and a cochlea
adapted for hearing the corresponding echoes. The craniofacial and inner ear
morphology of Oligocene odontocetes support a rapid (less than 5 Myr) early
evolution of echolocation. Although some cranial features in the stem
odontocetes Simocetus and Olympicetus suggest an ability to
generate ultrasonic sound, until now, the bony labyrinths of taxa of this grade
have not been investigated. Here, we use µCT to examine a petrosal of a taxon
with clear similarities to Olympicetus avitus. Measurements of the bony
labyrinth, when added to an extensive dataset of cetartiodactyls, resulted in
this specimen sharing a morphospace with stem whales, suggesting a transitional
inner ear. This discovery implies that either the lineage leading to this Olympicetus-like
taxon lost the ability to hear ultrasonic sound, or adaptations for ultrasonic
hearing evolved twice, once in xenorophids and again on the stem of the
odontocete crown group. We favour the latter interpretation as it matches a
well-documented convergence of craniofacial morphology between xenorophids and
extant odontocetes.
Comment:
This paper started out from the first completely prepared dolphin from the
James Goedert collection at the Mace Brown Museum of Natural History to have a
periotic- which we offered to Dr. Rachel Racicot to CT scan, just to see if
there was anything interesting. Then it turned out that this dolphin had none
of the hallmark features that are a requirement for echolocation - and based on
its phylogenetic position, requires either 1) a single loss of echolocation in
this lineage or 2) independent derivation or elaboration of echolocation within
xenorophids and main-line odontocetes. For various reasons we laid out, the
latter option is more likely - but awaits testing by study of other
simocetid-grade odontocetes. A surprising reward for four months of acid
preparation in a rubbermaid bin in our lab, and 5 minutes of scrubbing a week!

Ramasammy and Lauridsen. A new specimen of Ziphiidae (Cetacea,
Odontoceti) from the late Miocene of Denmark with morphological evidence for
suction feeding behaviour. Royal Society Open Science.
Link:
https://royalsocietypublishing.org/doi/10.1098/rsos.191347
Abstract:
A new fossil of Ziphiidae from the upper Miocene Gram Formation (ca
9.9–7.2 Ma) is described herein. Computed tomographic scanning of the specimen
was performed to visualize the mandibles and to obtain a three-dimensional
digital reconstruction. It possesses several characters of the derived
ziphiids, such as the dorsoventral thickening of the anterior process of the
periotic, the dorsoventral compression of the pars cochlearis and the short
unfused symphysis. The specimen cannot be identified beyond the family level,
because of the unusual nature of the preserved parts consisting of the
mandibles, earbones and postcranial remains. It differs from other ziphiid
species from the Gram Formation, Dagonodum mojnum, in its larger size
and the more derived morphology of its mandibles and earbones. Its long and
thickened stylohyal, combined with its reduced teeth, suggests that this new
specimen relied primarily on suction feeding. By contrast, the other ziphiid
species from the Gram Formation, D. mojnum, shows adaptations for a more
raptorial feeding strategy. Assuming the two species were coeval, their
co-occurrence at the same locality with two different feeding strategies, may
represent a case of niche separation. They may have hunted different types of
prey, thus avoiding direct competition for the same food resource.

Samonds et al. New Miocene sirenians from Nosy
Makamby, northwestern Madagascar. Journal of Vertebrate
Paleontology.
Link:
https://www.tandfonline.com/doi/abs/10.1080/02724634.2019.1570223
Abstract:
The near lack of vertebrate fossils from the Cenozoic of Madagascar has
left many of the details regarding the origin and evolution of the island’s
extant faunas unknown. However, recent fossil discoveries from Madagascar’s
nearshore marine deposits have begun to elucidate this mystery. These finds
include sharks, bony fish, turtles, crocodylians, a middle Eocene sirenian (Eotheroides
lambondrano), and the island’s first fossil dolphin. We report here at
least three (possibly four) different early (or possibly later) Miocene
dugongid sirenians recovered from the island
of Nosy Makamby, Mahajanga
Basin, northwestern Madagascar.
These include (1) a fragmentary braincase originally attributed to the genus Halitherium
but here reidentified as a previously named species known only from Libya (Rytiodus
heali; Dugonginae); (2) a newly named genus and species (Norosiren
zazavavindrano) interpreted as a primitive relative of Xenosiren
(Dugonginae); (3) a probable dugongine not yet identified with any known
species; and (4) a taxon reported here as Metaxytherium cf. krahuletzi
(Halitheriinae), the first Neogene halitheriine credibly reported from the
Indian Ocean basin. This pattern of shallow marine environments harboring
multispecies sirenian paleofaunas is seen elsewhere in the world, and these
three or four contemporaneous sirenians represent the first glimpse into Madagascar’s
sea cow diversity during the Miocene. This specific time period is a poorly
known and critical interval for interpreting Madagascar’s
past, and these specimens are potentially highly significant for reconstructing
sirenian evolutionary and biogeographic history. Surprisingly, this sirenian
fauna, so far, shares no genera with the roughly contemporaneous and relatively
nearby one from Kutch, western India.

Schipps et al. Borealodon
osedax, a new stem mysticete (Mammalia, Cetacea) from the
Oligocene of Washington State and its implications for fossil whale-fall
communities. Royal Society Open Science.
Link:
https://royalsocietypublishing.org/doi/10.1098/rsos.182168
Abstract:
Baleen whales (mysticetes) lack teeth as adults and instead filter feed
using keratinous baleen plates. They do not echolocate with ultrasonic frequencies
like toothed whales but are instead known for infrasonic acoustics. Both baleen
and infrasonic hearing are separately considered key innovations linked to
their gigantism, evolutionary success and ecological diversity. The earliest
mysticetes had teeth, and the phylogenetic position of many so-called toothed
mysticetes remains debated, including those belonging to the nominal taxonomic
groups Llanocetidae, Mammalodontidae and Aetiocetidae. Here, we report a new
stem mysticete, Borealodon osedax gen. et sp. nov., from the Oligocene
of Washington State, USA. Borealodon preserves multi-cusped teeth with
apical wear; microCT scans of the inner ear indicate that the minimum frequency
hearing limit of Borealodon was similar to mammalodontids. Borealodon
is not recovered within a monophyletic Mammalodontidae nor a monophyletic
Aetiocetidae; instead, it represents an unnamed lineage of stem Mysticeti,
adding to the diversity of stem mysticetes, especially across the
Rupelian–Chattian boundary. Furthermore, the presence of a putative
chemosynthetic bivalve along with Osedax, a bone-boring annelid, found
in association with the type specimen of Borealodon, offer more insights
into the evolution of deep-sea whale-fall communities.
Comments:
An interesting albeit fragmentary toothed mysticete - I first saw the specimen,
expertly prepared by the discoverer - James L. Goedert - in 2016. It was
originally presented as a mammalodontid in an SVP abstract a few years ago, an
affinity not supported in this paper (their analysis places it outside
Mammalodontidae and outside Aetiocetidae), so I suspect the name - Borealodon -
was a nod to Mammalodon but kept regardless of the affinity. More complete
fossils currently under study will shed light on the affinities of this poorly
understood toothed mysticete.
Serio et al. Macroevolution
of Toothed Whales Exceptional Relative Brain Size. Evolutionary Biology.
Link:
https://link.springer.com/article/10.1007%2Fs11692-019-09485-7
Abstract:
Toothed whales (Odontoceti, Cetacea) are well-known for their ability to
produce complex vocalizations, to use tools, to possess self-recognition, and
for their extreme behavioural plasticity. The toothed whale intelligence is
said to compete with that of primates, so does their extremely large brain to
body size ratio. Common explanations for the acquisition of such large brains
over the evolutionary time (encephalization) in toothed whales range from their
demanding, complex social lives, to their feeding habits, to echolocation. Yet,
several studies found no macroevolutionary trend in Odontoceti encephalization,
which casts doubts on its selective advantage. We applied a recently developed
phylogenetic comparative method to study macroevolutionary trends in relative
brain size (RBS) and brain size evolutionary rates in cetaceans, comparing
toothed whales to the other cetaceans and contrasting groups of species as
ascribed to different feeding categories. We found that cetaceans as a whole
followed a trend for increased encephalization over time, starting from
small-brained archaeocete ancestors. Toothed whales do not show this same trend
in RBS but have possessed larger RBS than any other cetacean ever since the
beginning of their existence. The rate of RBS evolution in Odontoceti is
significantly slower than in other Cetacea and slower than the rate of
Odontoceti body size evolution. These results suggest that toothed whales’
history is characterized by high and conservative relative encephalization.
Feeding lifestyle does not explain these patterns, while the appearance of
echolocation within stem group Odontoceti remains a viable candidate for them.

Shinmura
and Matsui. Life appearances of Desmostylia using a three-dimensional
computer graphics (3D CG). Fossils.
Link: http://www.palaeo-soc-japan.jp/publications/106_FrontPage_Shinmura_Matsui.pdf
Abstract: In Japanese (can't copy/paste).
Tanaka
and Taruno. The First Cetacean Record from the Osaka Group (Middle
Pleistocene, Quaternary) in Osaka, Japan.
Paleontological Research.
Link:
https://bioone.org/journals/Paleontological-Research/volume-23/issue-2/2018PR016/The-First-Cetacean-Record-from-the-Osaka-Group-Middle-Pleistocene/10.2517/2018PR016.short
Abstract:
A new partial skeleton consisting of a left mandible and five caudal
vertebrae, OMNH-QV 282 from the Osaka Group (Middle Pleistocene, about 0.3
million years ago) of Osaka City
is reported as the first cetacean record from the group. The skeleton is
identified as Balaenopteridae gen. et sp. indet. based on the combination of
mandibular characters, such as having a small mandibular foramen, reflected
neck in dorsal view and lack of a satellite process of the mandible. OMNH-QV
282 expands diversity for the local fauna, and also adds evidence for the
existence of large-sized balaenopterids from a poorly known epoch, the Middle
Pleistocene.
Tanaka
et al. A Large Fossil Baleen Whale from the Shikiya Formation (Early
Middle Miocene) of Wakayama, Japan.
Paleontological Research.
Link:
https://bioone.org/journals/Paleontological-Research/volume-23/issue-3/2018PR020/A-Large-Fossil-Baleen-Whale-from-the-Shikiya-Formation-Early/10.2517/2018PR020.short
Abstract:
A new large Chaeomysticeti indet., WMNH-Ge-1140240005 from the Shikiya
Formation of Kumano Group (early middle Miocene; about 16 to 15 Ma) of Wakayama,
Japan is
described here. It preserves a large rostrum (about 50 cm width at the base of
the rostrum), which has gently tapered lateral margins of the rostrum, narrow
mesorostral groove at the level of the narial fossa, and wide premaxillae and
maxillae. There are no diagnostic features pertaining to the specimen at the
family level, but it is comparable to “cetotheres” sensu lato such as Pelocetus
calvertensis and Diorocetus hiatus by having wide premaxillae that
occupy 1/3 width of the rostrum anterior to the narial fossa in dorsal view,
which implies that WMNH-Ge-1140240005 is a possible member of “cetotheres” sensu
lato. Its size is possibly between the two large species Pelocetus
calvertensis and Diorocetus hiatus of the early middle Miocene, and
larger than the reported middle Miocene mysticete specimens from Japan (“Diorocetus”
chichibuensis, “Diorocetus” shobarensis, Parietobalaena
sp. (SMNH-VeF-62)). The rostral width of WMNH-Ge-1140240005 suggests that this
animal was medium-sized compare to extant species, but the largest in the class
of baleen whales of its time.

Tanaka and Watanabe: An early and new member of
Balaenopteridae from the upper Miocene of Hokkaido, Japan. Journal of Systematic
Palaeontology.
Link:
https://www.tandfonline.com/doi/full/10.1080/14772019.2018.1532968
Abstract:
The family Balaenopteridae includes the modern blue whale, which is the
largest animal on Earth. Our knowledge of the early members of Balaenopteridae,
especially during the late Miocene and earlier periods, has recently increased,
but this family is still poorly understood. A fossil balaenopterid (including
the frontals, squamosals, parietals, vomer, pterygoid, basisphenoid,
basioccipital, exoccipitals, right periotic and presternum) from the lower part
of the Horokaoshirarika Formation, late Miocene (6.5–6.8 Ma) of Hokkaido,
Japan is named
as a new genus and species Miobalaenoptera numataensis, and is placed in
a phylogenetic context. As a result, Miobalaenoptera numataensis is
placed among the stem balaenopterids. The periotic of Miobalaenoptera
numataensis shows a deeper suprameatal fossa and a shorter caudal tympanic
process than extant species, incipient lateral tuberosity and a sharp
anteroposteriorly long ridge on the ventral side of the anterior process. Miobalaenoptera
numataensis adds information on the early morphology of the family
including the periotic.

Tanaka
et al. Crown beaked whale fossils from the Chepotsunai Formation (latest
Miocene) of Tomamae Town, Hokkaido, Japan. Palaeontologia Electronica.
Link:
https://palaeo-electronica.org/content/2019/2544-ziphiid-from-japan
Abstract:
In the last decades, our knowledge of ziphiid evolution has increased
dramatically. However, their periotic morphology is still poorly known. A
fossil ziphiid (TTM-1) including the periotic, bulla, isolated polydont teeth
and vertebrae from the Chepotsunai Formation (latest Miocene) of Tomamae Town,
Hokkaido, Japan, is identified as a member of a clade with crown ziphiids of
Bianucci et al. (2016) by having three periotic synapomorphies; a posteriorly
wide posterior process, transversely thick anterior process, and laterally
elongated lateral process. The specimen adds morphological information of the
periotic. Among the Ziphiidae from the stem to crown, the periotic morphologies
were changed to having a more robust anterior process, wider anterior bullar
facet and posterior process. The crown Ziphiidae shares a feature; enlarged
medial tubercle on the anterior process. Among the crown Ziphiidae, TTM-1 does not
have a swollen medial tubercle not like Tasmacetus, Nazcacetus and
others. This new morphological information might represent useful future
phylogenetic comparisons.

Taylor et al. - Isotopes from fossil coronulid barnacle shells record evidence of migration in multiple Pleistocene whale populations. Proceedings of the National Academy of Sciences.
Link: https://www.pnas.org/content/116/15/7377
Abstract: Migration is an integral feature of modern mysticete whale ecology, and the demands of migration may have played a key role in shaping mysticete evolutionary history. Constraining when migration became established and assessing how it has changed through time may yield valuable insight into the evolution of mysticete whales and the oceans in which they lived. However, there are currently few data which directly assess prehistoric mysticete migrations. Here we show that calcite δ18O profiles of two species of modern whale barnacles (coronulids) accurately reflect the known migration routes of their host whales. We then analyze well-preserved fossil coronulids from three different locations along the eastern Pacific coast, finding that δ18O profiles from these fossils exhibit trends and ranges similar to modern specimens. Our results demonstrate that migration is an ancient behavior within the humpback and gray whale lineages and that multiple Pleistocene populations were undertaking migrations of an extent similar to those of the present day.
Tsai and Chang. A right whale (Mysticeti,
Balaenidae) from the Pleistocene of Taiwan. Zoology Letters.
Link:
https://zoologicalletters.biomedcentral.com/articles/10.1186/s40851-019-0153-z
Abstract: Current
patterns of biological distribution result from the deep past. Of particular
interest, some closely related species appear at high latitudes of both
hemispheres, but not in between, a pattern known as antitropical distribution.
However, the timing, pathway, and drivers of antitropical distributions remain
mostly unknown. Here we describe a new fossil, a left tympanic bulla (part of
the ear bones), from the Middle/Late Pleistocene (0.78–0.01 mya, but not
excluding the possibility of Holocene in age, as the specimen was dredged from
the sea bottom and the geological horizon remains uncertain) of Taiwan.
The tympanic bulla is diagnostic in baleen whales, and this specimen shows
morphological features that are identical to extant Eubalaena,
including: relatively large size (the anteroposterior length is 117 mm);
rectangular outline in medial view; short anterior lobe, judging from the
remaining of the lateral furrow; squared anterior margin; prominent transverse
crease on the involucrum; transversely compressed in anterior view;
well-developed and rounded outer lip; and parallel involucral and main ridges.
Although incomplete, the morphological characters and overall similarity to
extant Eubalaena allow a reliable taxonomic assignment to Eubalaena
sp. The occurrence of a Pleistocene Eubalaena on the southern margin of
the western North Pacific is the first balaenid fossil evidence indicative of
the biotic interchange between two hemispheres leading to the origin of
antitropical distribution in the Pleistocene; alternatively, this specimen
might merely represent an extra-limital record of the North Pacific Eubalaena.
Furthermore, this find suggests that the Eubalaena interchange, being
one of the largest species displaying antitropical distribution pairs in the
history of life, likely took place along the western Pacific. Notably, this
does not preclude the Eubalaena interchange from other routes, such as
the eastern Pacific or the Atlantic Ocean, and future
finds should test the scenario for the biotic interchange between Northern and
Southern Hemispheres of Eubalaena.
Comments: CH Tsai and I have
already published a couple of papers on isolated yet quite critical Pleistocene
mysticete bullae from northern California - so it was nice to see him
continuing the practice on the opposite side of the Pacific! Let's just say
there are many more surprising fossils yet to be reported from the Penghu
Channel and elsewhere in Taiwan. Cheers to Tsai for
reinvigorating the study of marine mammal evolution in Taiwan.
Valenzuela-Toro and Pyenson. What do we know about the fossil record of pinnipeds? A historiographical
investigation. Royal Society Open Science.
Link:
https://royalsocietypublishing.org/doi/10.1098/rsos.191394
Abstract:
The fossil record of pinnipeds (seals, fur seals and walruses) is
globally distributed, spanning from the late Oligocene to the Holocene. This
record shows a complex evolutionary history that could not otherwise be
inferred from their extant relatives, including multiple radiations and
iterative ecomorphological specializations among different lineages, many of
which are extinct. The fossil record of pinnipeds is not uniformly represented
in space and time, however, leaving some gaps in our knowledge. We performed a historiographical
investigation of the published fossil record of pinnipeds based on the
information available in the Paleobiology Database, with the aim to broadly
characterize and evaluate it from a taxonomic, geographical and temporal
perspective. We identified major trends, strengths and weaknesses of the
pinniped fossil record, including potential biases that may affect our
interpretations. We found that 39% of the record corresponds to extant taxa,
which are essentially from the Pleistocene and Holocene. There is a larger
record from the Northern Hemisphere, suggesting biases in sampling and
collection effort. The record is not strongly biased by sedimentary outcrop
bias. Specifically, for extinct species, nearly half of them are represented by
a single occurrence and a large proportion have type specimens consisting of
single isolated postcranial elements. While the pinniped fossil record may have
adequate temporal and taxonomic coverage, it has a strong geographical bias and
its comparability is hindered by the incompleteness of type specimens. These
results should be taken into account when addressing patterns of their past
diversity, evolutionary history and paleoecology.

Vautrin et al. From limb
to fin: an Eocene protocetid forelimb from Senegal sheds
new light on the early locomotor evolution of cetaceans. Palaeontology.
Link:
https://onlinelibrary.wiley.com/doi/full/10.1111/pala.12442
Abstract:
Cetaceans constitute a textbook example of the secondary adaptation of
tetrapods to aquatic life. This major event in the evolutionary history of
mammals is often linked in the literature to the limb‐to‐fin
transition. Paradoxically, limb bones are scarce in the fossil record of early
cetaceans, and the transition from a limb‐adapted morphology for an
amphibious life in shallow water to a fin‐adapted morphology for a
pelagic lifestyle remains poorly documented. Here, we describe new protocetid
remains from the upper Lutetian of Senegal, including a nearly complete
articulated forelimb. A cladistic analysis including 24 taxa and 137
morphological characters recovers the new African specimen close to Carolinacetus.
It also confirms that cetacean dispersal to the New World
was not the result of a single colonization event. A 3D model of the forelimb
was reconstructed. Anatomical comparisons suggest that it is unlikely that the
Senegalese forelimb was used as a rigid pectoral flipper for steering as in
basilosaurids and modern cetaceans. Instead, we suggest that the hand was
actively used during swimming. This challenges previous reconstructions of
protocetids as mainly foot‐powered swimmers, and suggests that swimming
specializations of early cetaceans were probably more diverse than previously
considered.

Velez-Juarbe and Valenzuela-Toro. Oldest record of monk seals from the North Pacific and biogeographic
implications. Biology Letters.
Link:
https://royalsocietypublishing.org/doi/10.1098/rsbl.2019.0108
Abstract:
True seals (crown Phocidae) originated during the late Oligocene–early
Miocene (approx. 27–20 Ma) in the North Atlantic/Mediterranean region, with
later (middle Miocene, approx. 16–11 Ma) dispersal events to the South
Atlantic and South Pacific. Contrasting with other pinnipeds, the
fossil record of phocids from the North Pacific region is scarce and restricted
to the Pleistocene. Here we present the oldest fossil record of crown phocids,
monachines (monk seals), from the North Pacific region. The specimens were
collected from the upper Monterey Formation in Southern California
and are dated to 8.5–7.1 Ma, predating the previously oldest known record by at
least 7 Ma. This record provides new insights into the early biogeographic
history of phocids in the North Pacific and is consistent with a northward
dispersal of monk seals (monachines), which has been recognized for other
groups of marine mammals. Alternatively, this finding may correspond with a
westward dispersal through the Central American Seaway of some ancestor of the
Hawaiian monk seal. This record increases the taxonomic richness of the Monterey
pinniped assemblage to five taxa, making it a fairly diverse fossil assemblage,
but also constitutes the oldest record of sympatry among all three extant
pinniped crown clades.
Comments: This surprising discovery reports a couple of very
fragmentary but distinctive teeth of monachine seals from a bonebed in the
Monterey Formation. These teeth would be typical for Peru or even here in South
Carolina - but the story up until this (told even in my own papers) was that
phocid seals did not arrive in the eastern North Pacific until the middle
Pleistocene, with the dispersal of harbor seals through the Arctic and elephant
seals northward across the equator from the southern ocean. These rare
specimens, and the lack of skulls or obvious monachine limb bones, suggests to
me that these may represent vagrants rather than a resident
breeding population of monk seals.
Velez-Juarbe and Wood. An early Miocene dugongine (Sirenia: Dugongidae) from Panama. Journal of Vertebrate Paleontology.
Link: https://www.tandfonline.com/doi/full/10.1080/02724634.2018.1511799
Abstract: Herein, we describe a new early Miocene dugongine from marine deposits of the Culebra Cut (Gaillard Cut) of the Panama Canal. The new taxon, Culebratherium alemani, gen. et sp. nov., represents one of the few records of late Aquitanian–early Burdigalian sirenians and the oldest sirenian from Central America. A phylogenetic analysis places Culebratherium in a clade with Dioplotherium cf. D. allisoni (Miocene of Brazil), Dioplotherium allisoni (Miocene of Baja California Sur, Mexico, and California, U.S.A.), and Dioplotherium sp. (Pliocene of Yucatan, Mexico). Similar to these taxa, Culebratherium is characterized by the presence of large incisor tusks, a premaxillary symphysis without a boss, a premaxilla-frontal suture forming a butt joint, and a moderately downturned rostrum. In addition, Culebratherium exhibits prominent occipital-cervical attachment sites for enlarged neck musculature. These features taken together are interpreted as adaptations for uprooting large, deeply buried seagrass rhizomes. Other dugongines with similar, yet convergent, dental and facial adaptations are known from earlier or coeval deposits in Puerto Rico, Florida, South Carolina, California, Baja California Sur, Brazil, and India and were constituents of sympatric paleocommunities of sirenians. Only fragmentary evidence of a second smaller and unidentifiable sirenian species is known from the Culebra Formation, but future discoveries may reveal a similar sympatric paleocommunity during the early Miocene of Panama. Finally, we used the results of the phylogenetic analysis to propose the new clade Pan-Sirenia as the most inclusive group consisting of stem and crown groups and redefine the Sirenia, Dugongidae, and Dugonginae clades.
Voss et al. - Stomach contents of the archaeocete Basilosaurus isis:
Apex predator in oceans of the late Eocene. PLoS One.
Link:
https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0209021
Abstract: Apex
predators live at the top of an ecological pyramid, preying on animals in the
pyramid below and normally immune from predation themselves. Apex predators are
often, but not always, the largest animals of their kind. The living killer
whale Orcinus orca is an apex predator in modern world oceans.
Here we focus on an earlier apex predator, the late Eocene archaeocete Basilosaurus
isis from Wadi Al Hitan in Egypt, and show from stomach contents
that it fed on smaller whales (juvenile Dorudon atrox) and large
fishes (Pycnodus mokattamensis). Our observations, the first direct
evidence of diet in Basilosaurus isis, confirm a predator-prey
relationship of the two most frequently found fossil whales in Wadi
Al-Hitan, B. isis and D. atrox.
This extends our understanding of their paleoecology. Late Eocene Basilosaurus
isis, late Miocene Livyatan melvillei, and modern Orcinus
orca are three marine apex predators known from relatively short
intervals of time. Little is known about whales as apex predators through much
of the Cenozoic era, and whales as apex predators deserve more attention than
they have received.
Comments:
Earlier studies by Julia Fahlke demonstrated bite marks on juvenile Dorudon
atrox skulls, presumed to be made by Basilosaurus isis. This awesome discovery
reports the gut contents of Basilosaurus - including Dorudon bones and teeth,
and fish/shark remains. Gut contents were reported for Dorudon atrox as well by
Mark Uhen in his 2004 monograph, and fish bones were reported as gut contents
in Basilosaurus isis ages ago in a
still-unpublished SVP abstract.
Werth et al. Enamel Microstructure in Cetacea: a Case Study in Evolutionary Loss of Complexity. Journal of Mammalian Evolution.
Link: https://link.springer.com/article/10.1007/s10914-019-09484-7
Abstract: Enamel microstructure of 34 species in eight odontocete families is described and categorized according to structural complexity (from no or prismless enamel to highly arranged enamel in decussating layers), then correlated with parameters of ecology, life history, and occlusal function. Overall, more complex dental structure in extant and extinct cetaceans is associated with smaller, more numerous teeth in taxa that bite or grasp smaller, harder prey with longer, narrower jaws and have more oral processing. Enamel complexity loosely correlates with mechanical properties, but measurement of compressive strength is complicated by the presence of cementum overlying or in place of crown enamel. Given the presence of Hunter-Schreger bands and other indices of high microstructural complexity in archaic cetaceans (with presumed plesiomorphic characters), the absence of complex enamel, as well as more generally of gross dental form, signals a loss of complexity in various cetacean lineages. Nonetheless, it is difficult to draw robust or reliable inferences regarding either cetacean phylogeny or dental function given the pattern of presumed loss of tooth complexity, and the confounding presence of numerous exceptions (taxa with complex teeth despite limited use, or with thin, weakly developed enamel and soft, simple teeth despite high occlusal function). Although retention of a complex feature implies its continued adaptive value, and thus selection for the underlying genetic and developmental bases for that complexity, loss of complexity can simply follow absence of function. Loss of complex traits presumably relates to accumulated mutations and economic efficiency, but unfolds with no distinct pattern of distribution.
Woolley et al. Microanatomy and histology of
bone pathologies of extant and extinct phocid seals. Historical Biology.
Link: https://www.tandfonline.com/doi/abs/10.1080/08912963.2019.1689238
Abstract: This study investigates three presumed fractured
phocid seal bones: An isolated metapodial and an ulna belonging to different
individuals of the extinct phocid, Homiphoca capensis, from Langebaanweg
and a mandible of a juvenile elephant seal (Mirounga leonina), which was
included to assess the validity of the assumption that changes to bones caused
by fractures are consistent across extant and extinct members of the same
groups. The bones were studied using a multi-method approach, including gross
morphology, microcomputed tomography (micro-CT) and histology. Micro-CT showed
that the metapodial was not fractured and information drawn from other analyses
suggested that the pathology was an osteosarcoma. Histology of normal and
fractured regions of the mandible and ulna permitted an estimate about the
fracture healing stage, and showed the bone tissue at the fracture sites to be
histologically similar. A birth line found on the lateral surface of the
elephant seal mandible emphasised its young age and marked the first example of
a birth line in a bone of a semi-aquatic mammal. A large scope of information
was obtained using this multi-method approach, which could permit insight into
the life events and lifestyles of modern and extinct individuals, such as H.
capensis.