
Welcome to the fifth annual post chronicling the year's publications on the marine mammal fossil record. It's only a couple weeks into January, so I'm doing pretty good this year. Note that I include anything that was published first in 2016 - some of these already have 2017 publication dates as they were published "Online Early" in 2016 but assigned final volume/issue/page numbers since Jan 1. Because I do this, I do NOT transfer summaries from articles published online in 2015 but published in print in 2016 from last year's post to this one; I don't have the time, and you can just relax.
The usual disclaimer applies - I may have forgotten something. If so, leave a comment, send me an email, or facebook message - and I'll be sure to correct it. Note that Blogger has a nasty habit of deleting spaces and de-italicizing anything within hyperlinks, and I've tried re-adding spaces but won't bother with italicizing anything in the hyperlinks. I do this in my spare time, so don't you dare complain to me about it.

The fossil record of marine mammals is heavily biased toward
the northern hemisphere - making localities like New Zealand (where I did my
Ph.D.), Australia, Antarctica, and especially South America ripe targets for
paleontological exploration. Recent efforts have yielded the first archaeocetes
from South America, one of the only Aquitanian (earliest Miocene) marine mammal
assemblages anywhere on earth, and the embarrassingly productive regions of the
Pisco basin (Peru) and the Bahia Inglesa area (Chile). Marine mammals are today
linked to cold water temperatures and high primary productivity - making good
marine mammal watching (typically) latitudinally constrained. Similar biases
affect the fossil record of marine mammals - marine mammals are geologically
young (Ischyrorhynchus vanbenedeni, a longirostrine
relative of the modern Amazon river dolphin Inia).
Recent analyses suggest Ischyrorhynchus may actually be a platanistid -
and the new rostra referred to this taxon is really reminiscent of Platanista
in some regards. A scrappy iniid skull is also described, as well as a pair of
vertebrae referred to Zarhachis flagellator, a species known only
from the Chesapeake Group of the eastern USA. Other remains include a couple of
partial baleen whale skulls. The authors suggest that the river dolphin Saurocetes
may be a synonym of Ischyrorhynchus, and highlight the possibility of a
south American record of Zarhachis - which awaits further field study.

Multiple groups of marine mammals have made the plunge
(excuse the pun) and returned to life in the sea. At least two of these - the
sea cows and cetaceans - have become so modified for aquatic life that they
cannot support their weight on land. Others, like pinnipeds and sea otters,
return to land to mate or for rest. While modern animals can simply be observed
by someone with a notebook and binoculars, fossilized marine mammals have been
dead for a long time and don't exactly move around a whole lot - so paleontologists
interested in the evolution of secondarily aquatic tetrapods have to examine
anatomical (chiefly, skeletal, owing to the nature of the fossil record)
evidence. Much of the research on aquatic habits have focused on fore/hindlimb
functional morphology and flexibility of the vertebral column; other important
studies have examined the histology of postcranial bones as increases in bone
density often happen early in semiaquatic lineages. However, few - if any -
studies have examined changes in ribs (aside from chopping them up for
histology, of course!). This study alleviates this problem by compiling a
dataset of rib thickness measurements, intended to serve as a rough proxy of
bending strength - the idea being that fully aquatic species which never return
to land would have ribs that would break under the animal's body weight if it
laid down on a hard surface out of water (as often happens with stranded whales
and dolphins). They found that, unsurprisingly, all modern cetaceans have
relatively weak ribs which cannot support their weight on land, and that
pinnipeds and otters do have strong ribs (which matches modern observations).
Surprisingly, the early semiaquatic archaeocete whale Ambulocetus may
not have been able to support its weight on land without breaking its ribs -
surprising, and suggestive that Ambulocetus was obligately aquatic.
Further surprising are their findings of desmostylians - Desmostylus
could support its weight on land, but Paleoparadoxia and Neoparadoxia
could not. This study needs to be tested using CT data, but as simple as the
current study is it is provocative and certain to generate some interesting
future discussion.

Sirenians are relatively common in the fossil record as far
as marine mammals are concerned and include two major families: the
Trichechidae (manatees) and Dugongidae (dugongs and sea cows); the latter is
quite diverse, and includes a host of species which managed to disperse worldwide
including the modern Dugong and the recently extinct Hydrodamalis
gigas (Steller's sea cow). Dugongids diversified early in the North
Atlantic and Tethys, later dispersing into the Pacific in the early Miocene.
This study reports a new species of Prototherium, Prototherium
ausetanum, from the middle Eocene of Spain. The species is characterized by
a rather elongate and narrow cranium and a rostrum that is not deflected much.
Cladistic analysis links this species with Eotheroides aegyptiacum, and
suggests that either Prototherium may not be monophyletic or perhaps
that the cladistic matrix needs work; similarly, Halitherium and Eotheroides
(other early dugongids) were also found to be problematic genera.

Remingtonocetids are among the smallest of the archaeocetes,
previously known from the middle Eocene of Indo-Pakistan. Remingtonocetids are
unique in that they are one of the only groups of archaocetes that likely forms
an actual clade - pakicetids, ambulocetids, protocetids, and basilosaurids (in
order of increasing aquatic adaptations and closeness on the tree to modern
cetaceans) commonly plot out as paraphyletic 'families'. Remingtonocetids are
relatively small and otter-like in body form with relatively shorter legs than
the decidedly more terrestrial long-legged pakicetids and powerful tails, yet
possess a greatly elongated rostrum, an enormous saggittal crest, and dorsally
placed orbits, not unlike a crocodilian in general form. Indeed,
remingtonocetids have been inferred as ambush predators similar to
crocodilians, and I colloquially refer to them as "furry crocs" or
"otter-crocs". This new study by Ryan Bebej and others reports a new
remintonocetid from the middle Eocene of Egypt - extending the range of this
clade from the north central Tethys Sea
(Indo Pakistan)
a bit further to the southwestern Tethys (north Africa). The new species Rayanistes
afer lacks skull parts, and so this was only possible owing to earlier work
describing more complete skeletons of Remingtonocetus - naturally
building upon some of Bebej's Ph.D. work. The holotype consists of a pelvis and
sacral vertebrae, a caudal vertebra, and a well-preserved femur - one of the
only remingtonocetid femora known. The pelvis exhibits a greatly enlarged ilium
and the femur is more robust than Remingtonocetus, indicating greater
musculature involved in the hindlimb power stroke; the hip socket appears to
have permitted greater range of movement. Additionally the vertebral column
exhibits features indicating it was more 'passively flexible' than Remingtonocetus,
suggesting more efficiant swimming and paddling than in Remingtonocetus.
The discovery of a remingtonocetid in Egypt 1) indicates that remingtonocetids
had a greater capacity for dispersal than previously assumed (protocetids were
assumed to be the first cetaceans to leave Indo-Pakistan) and 2) raises the
possibility that other early semiaquatic archaeocetes (e.g. ambulocetids) may
be found in Egypt as well, or similarly that remingtonocetids may be found
further yet a field - which would prompt reevaluation of early whale evolution.

Baleen whales are of course one of my favorite research
subjects - I was a bit sick of them after my Ph.D. on eomysticetids, but that's
finally beginning to wear off. Quite a lot has been published in the last
decade: the description of Janjucetus and redescription of Mammalodon,
the discovery that aetiocetids may have had teeth and baleen, the discovery and
naming of numerous eomysticetids from New Zealand and Japan, discovery of
aberrant early chaeomysticetes (Whakakai, Horopeta, and the
possible eomysticetid Sitsqwayk - see below), the discovery that
eomysticetids may have retained teeth, major advances in mysticete phylogeny
including the provocative hypothesis that the pygmy right whale is related to
cetotheriids, the identification of genetic processes leading to tooth loss,
clarification of mysticete hearing and olfaction, and the "cetothere
revolution". This review article by Annalisa Berta and others covers all
recent advances regarding the dramatic evolution of the feeding apparatus of
baleen whales, and is a fine starting point for new researchers to become
acquainted with baleen whale evolution (heaven forbid baleen whales attract any
additional research attention...). Unfortunately this paper came out just too
late to incorporate information from the Marx et al. study (see below) on evidence
of suction feeding in aetiocetids.

A majority of the beaked whale fossil record consists of
partial crania dredged from seafloor deposits without age information. The
Pisco Formation of Peru is one of the few continental localities where
well-preserved partial skeletons of ziphiids are preserved in abundance. This
new study reports two new genera and species of ziphiids from the Pisco
Formation: Chavinziphius maxillocristatus and Chimuziphius
coloradensis. Chavinziphius has a facial region of the skull
convergent with giant beaked whales Berardius spp., yet retains a
functional homodont dentition like most odontocetes (most extant ziphiids only
retain tusks). Chimuziphius does not resemble modern species but exhibits
a mesorostral canal roofed over by the premaxillae, and triangular nasal bones.
Two additional ziphiids from the Pisco indicate the presence of five species
from the mid-late Miocene – they do not represent either genus named in this
paper, nor are they referable to Messapicetus gregarius; this assemblage
represents an unusually diverse ziphiid fauna. This study includes a new
cladistic analysis with a new phylogenetic hypothesis – many extinct and quite
bizarre ziphiids form a clade with the longirostrine and polydont ziphiid Messapicetus,
called the “Messapicetus clade” (completely extinct). This new
phylogenetic framework permits a reevaluation of character evolution in beaked
whales. Both crown Ziphiidae and the Messapicetus clade independently trend
towards reduction of dentition, increasing bone density of the facial region,
elevation of the vertex, and increasing body size.
Many modern whales and dolphins occur in both the southern
and northern hemispheres but rarely cross the equator - resulting in two
semi-disparate populations that are somewhat or completely reproductively
isolated. Warmer temperatures and lower productivity at the equator make it difficult
for individual whales and dolphins to cross because of overheating and lack of
food - though some larger baleen whales evidently do cross some of the time
(e.g. blue whales) and there are cold-water areas of upwelling that permit
intermittent cooling (Humboldt current, eastern equatorial Pacific). This
pattern is called antitropicality, and it represents a classic example of
allopatric speciation (speciation occurring in different areas); it's resulted
in restricting gene flow between two populations of the same species (e.g. fin
whales, blue whales, killer whales), and splitting of a single genus into two
species (e.g. right whales, bottlenose whales, giant beaked whales). We have
evidence of antitropical distributions in the fossil record of marine mammals
such as the porpoise Piscolithax, known from the late Miocene of
California and Peru. This study reports new specimens of the longirostrine
beaked whale Messapicetus longirostris from Italy, confirming
species-level separation between this and Messapicetus gregarius from
Peru. This relationship suggests that these ziphiids were an antitropical
species pair, inhabiting the Mediterranean and eastern South Pacific; no
remains of Messapicetus have yet been recorded from the eastern North Pacific,
where beaked whale fossils are relatively rare.

Balaenopterids (aka rorquals) are the baleen whales with
throat pleats, such as minke, fin, humpback, and blue whales; these are perhaps
my favorite group of cetaceans. Balaenopterids are quite common in late Miocene
and Pliocene marine sediments worldwide, but until the last decade or so have
remained very poorly known; the fossil record of balaenopterids has been
plagued by poor taxonomic practices dating from the 19th century to the 1970s.
P.J. Van Beneden described a number of Pliocene balaenopterids from Belgium,
but designated no holotypes and furthermore assembled chimera skeletons
composed of isolated, non-associated bones based on preconceived notions of
what each species should look like. Using modern diversity as a guide, he named
different species (all probably nomina dubia) for their sizes supposedly
mirroring modern species: Balaenoptera musculus ("B.
sibbaldina", "B. musculoides"), Megaptera novaeangliae
("Megapteropsis robusta"), and B. acutorostrata ("B.
rostratella") and further re-named some of these species. In Italy,
individual specimens have been designated too many names, and all Pliocene
balaenopterids were once rolled into modern Balaenoptera acutorstrata (none
actually belong to minke whale-like species). This taxonomic paralysis has
plagued the study of fossil balaenopterids, and crania associated with
mandibles and earbones are needed to evaluate old names (or, throw them out
entirely). This new study reports a fragmentary but informative skull from the
Pliocene of Belgium that represents a new genus, named Fragilicetus velponi.
It appears to be different than all of Van Beneden's figured material
(taxonomic problems aside), and anatomically is relatively primitive in
comparison to other balaenopterids. Most critically, this taxon exhibits a
suite of features that are intermediate between balaenopterids and gray whales
(Eschrichtiidae). Gray whales and rorquals are known to be closely related, but
whether gray whales are sister to or included within the balaenopteridae is a
contentious issue as the modern species are very divergent in morphology.
Fossils like Fragilicetus suggest that the earliest crown Balaenopteroidea may
have exhibited features of both families, with later diverging members of each
clade evolving gray whale and rorqual features more and more until the present.

Modern river dolphins were formerly grouped into a single
group, the Platanistoidea, based on shared primitive features (e.g. long,
narrow rostra). Later morphological and eventually molecular studies revealed
that these modern dolphins (Inia, Amazon River dolphin; Lipotes,
Yangtze River dolphin; Pontoporia, Franciscana; and Platanista,
Indus/Ganges River dolphins) belong to different parts of the odontocete tree
and thus constitute a paraphyletic grouping. A second hypothesis, first
promoted by Christian de Muizon, proposed a new Platanistoidea that included
extant Platanista and fossil relatives – which have variably included
extinct platanistids, the squalodelphinids, allodelphinids, and in some
studies, waipatiids and squalodontids. This study came on the heels of a
monographic reevaluation of the Allodelphinidae – an Oligo-Miocene clade of
longirostrine dolphins from the North Pacific (see Kimura and Barnes, 2016,
below) – including Allodelphis, Goedertius, Ninjadelphis,
and Zarhinocetus. This new paper reports an additional genus and species
of allodelphinid, Arktocara yakataga, from the “middle” Oligocene of
southern Alaska. This new small-bodied odontocete is known only by a braincase,
and amongst the Allodelphinidae it is most similar to Allodelphis
from the Jewett Sand. This study critically includes the first cladistic
analysis including any allodelphinids, which confirmed the monophyly of the
group and position within the Platanistoidea sister to a
squalodelphinid-platanistid clade. Arktocara represents one of the
earliest known crown odontocetes (if platanistoid monophyly is accepted; larger
cladistic analyses indicate this group may be paraphyletic), though part of
this relates to the rather poor age control for this species (29-25 Ma, late
Rupelian to late Chattian).

The entire world's fossil record of true sea otters
(excluding Holocene age specimens) could probably fit inside of a briefcase.
Scrappy fossils of Enhydra have been reported from various localities of
supposed Pliocene and Pleistocene age along the California coast, Arctic coast
of Alaska, and England. Other giant otters include Enhydriodon from
Europe, Africa, and Asia, and Enhydritherium from the Pliocene of
California and Florida. In the 1960s-1970s, supposed Pliocene age of some
specimens of Enhydra from Los Angeles, coupled with the occurrence of
more primitive giant otter specimens (Enhydritherium), led early
researchers to propose that sea otters evolved in place in the eastern North
Pacific. Many discussions have revolved around the age of Enhydra reevei
from England – which is an isolated tooth nearly identical to modern sea
otters, and approximately 2 Ma in age. Because few anatomically informative Enhydra
specimens have been found, sea otter evolution and biogeography have hinged
upon the identification and age of individual occurrences. A new specimen of Enhydra
collected by a buddy of mine consists of a nearly complete femur from the
middle Pleistocene Merced Formation near San Francisco – a rock unit that is
dated exceedingly well. Available dates indicate an age of approximately
620-670 Ka – a fifty thousand year age control, which is staggeringly narrow.
This specimen prompted a reevaluation of the ages of various sea otter
occurrences in the Pacific. As it happens, all supposed Pliocene and early
Pleistocene specimens of Enhydra from the Pacific are actually much younger,
and either middle or late Pleistocene in age, and none older than 700 Ka – with
this femur being the oldest accurately dated specimen in the Pacific. Specimens
from Alaska and England
are early Pleistocene (~1-2 Ma) and predate the Pacific record – negating prior
arguments which sought to invalidate the role of Enhydra reevei. The
available record of Enhydra fossils thus support a North
Atlantic or even Arctic origin of true sea otters, followed by
dispersal through the Arctic into the Pacific –
indicating that sea otters are recent invaders.

The fossil record of pinnipeds is heavily biased toward the
northern hemisphere – and the north Pacific record is particularly densely
sampled. One of the most famous north Pacific pinnipeds is the smelly,
gigantic, bizarre, violent, and utterly charismatic elephant seal (Mirounga
angustirostris). It's one of only two temperate latitude true seals in the
eastern North Pacific, aside from the harbor seal. Whereas the closest
relatives of harbor seals all live in the North Pacific, North Atlantic, and
Arctic, the closest relatives of northern elephant seals (southern elephant
seal, Mirounga leonina, and other lobodontines) are all in the southern
ocean – making their evolutionary history a bit of a puzzle. Modern
biogeography suggests that the modern northern elephant seal originated in the
southern hemisphere with other lobodontines (antarctic seals). Unfortunately,
the fossil record of elephant seals is a bit crap; fragmentary specimens
indicate the presence of modern species in the mid-late Pleistocene of
California and Chile – indicating that the north-south split between the
species is at least a few hundred thousand years old, but not giving us a
better idea of which direction. Morphological studies initially proposed that
the northern elephant seal is more primitive, indicating a North Pacific
origin; however, phocid seals are unknown in pre-Pleistocene rocks in the North
Pacific, and the sample is densely sampled enough in my opinion that we can
confidently say that true seals are recent invaders. A “new” fossil seal – a
fragmentary skull collected in the 1920s and informally known as the “Waipunga
seal” was studied by Dr. J.A. Berry, a zoologist in NZ but never formally
published; other specimens of his were eventually published by the late Dr.
Judith King, one of the foremost pinnipedologists of the 20th
century. Morgan Churchill and I studied this specimen together when he visited
U. Otago for an EAPSI grant. This fragmentary skull is about the size of a
modern harbor seal, but evidently had a prenarial shelf and likely a small
proboscis like an elephant seal, but primitively retained double rooted teeth
with cusps. The bulbous crowns are shared uniquely with modern Mirounga,
but the more primitive features and incomplete preservation precluded us from
identifying it to the genus level, though identification to the Miroungini (a
group within the lobodontines). The age of this specimen is late Pliocene,
predating all fossils of Mirounga. This specimen thus represents some of
the first fossil evidence for a southern hemisphere origin of elephant seals,
sometime in the Pliocene – followed by a Pleistocene dispersal to the northern
Pacific. The specimen further suggests that Mirounga evolved from a
fairly small-bodied ancestor, perhaps only the size of a gray seal – and
evolved gigantism within the last 2 million years.

This study is the last major chapter from my Ph.D. thesis
and reports the new genus and species Matapanui waihao, the
geochronologically oldest eomysticetid baleen whale from New Zealand.
Eomysticetids are the earliest obligate filter-feeding baleen whales, were
mostly toothless, but likely had a few non-functional teeth in the front of the
jaws. Matapanui is known by several partial skulls with well-preserved
earbones, and is somewhat older than other NZ eomysticetids like Tokarahia,
Tohoraata, and Waharoa. Matapanui has some features reminiscent
of northern hemisphere eomysticetids, but cladistic analysis indicates it is
sister to the Tokarahia-Tohoraata-Waharoa clade –
indicating that most New Zealand eomysticetids form a clade. Whereas Matapanui
is not the most completely known eomysticetid, but it is known from numerous
referred specimens that reveal aspects of ontogenetic and intraspecific
variation. One of the specimens, a partial braincase, is from a large adult and
exhibits some features that are a bit of a departure from other eomysticetid
braincase anatomy including thickened nuchal crests, a greatly inflated
exoccipital, and a convex (rather than triangular) supraoccipital shield. At
approximately 27 Ma, Matapanui represents one of the oldest described
chaeomysticetes. In addition to description and cladistic analysis, I decided
to include within this paper a taxonomic review of the Eomysticetidae,
essentially summarizing all published literature on the clade. This includes
reporting updated geochronologic ranges for all named eomysticetids, and
formally recognizing Micromysticetus spp. as eomysticetids, and
suggesting that Cetotheriopsis lintianus – the type species of the
“Cetotheriopsidae” - is too incomplete to be the foundation of a family-level
clade, but has some features consistent with eomysticetids. Lastly, I'll add
that we originally published this as Matapa waihao; Matapa means
“flat face” in Maori. With prior names I found no evidence of occupied names,
given the unique structure of Polynesian words. However, Matapa is a bit
shorter and more simple than Tokarahia or Tohoraata, and
unbeknownst to me, Matapa had already been used as the genus name for a
butterfly in Sri Lanka
– which I was informed of within a day or so of the paper being published. So,
we went ahead and proposed the new genus name, Matapanui – meaning “big
flat face” in Maori.

Basilosaurid whales are the first truly pelagic whales to
have evolved - protocetids, though capable of crossing ocean basins, were
evidently still tied to land for reproduction similar to seals. Protocetids are
essentially unknown from the Pacific basin, though they escaped the closing
Tethys sea into the North Atlantic; a possible
protocetid from Peru
has affinities with Basilotritus, a basilosaurid formerly thought to be
a protocetid. Archaeocetes in general are quite rare from the southern
hemisphere - their record includes some well-preserved skeletons from Peru, a
partial skull and various teeth from New Zealand, and some scraps from
Antarctica. A number of kekenodontids are known as well from NZ -
late-surviving Oligocene archaeocetes. New material from the mid-late Eocene La
Meseta Formation of Seymoura Island, Antarctica, reported in this study by
Monica Buono and others significantly expands the record of southern
archaeocetes, and includes some nice mandibles as well as isolated teeth and a
nice pelvis. The fossils all represent basilosaurids, though none of the
material is complete enough to identify to the genus level. This material
supplements the scrappier remains reported by earlier researchers, and
critically none of the material appears to represent Neoceti - the
geochronologically earliest (but not necessarily most primitive) mysticete, Llanocetus
denticrenatus, originates from about the Eocene-Oligocene boundary about
100 meters up section in the La Meseta from some of these specimens. New
(albeit fragmentary) specimens of Llanocetus have been discovered by
this same team.

Fossils of sea cows (Sirenia) are rather common owing to
their dense bones that are resistant to fragmentation and abrasion. Like other
marine vertebrates, sirenians are typically discovered during weathering of
fossiliferous rocks exposed at the earth's surface. It's generally rare to find
anything during digging if no bones are exposed at the surface - a common mistake
in movies and TV shows. On occasion the improbable does happen, and this study
reports an isolated rib of a dugongid sea cow from the Floresta Calcarenites
Formation of southern Italy, recovered during sawing and polishing of
decorative tile slabs. Ribs are not identifiable to the genus or species level,
but it does extend the range of dugongids in southern Italy back to the
earliest middle Miocene or latest early Miocene. Additionally, the specimen is
absolutely beautiful. Similar finds include the protocetid Aegyptocetus
tarfa, recovered in Eocene Egyptian limestone, and a sea cow (Prototherium,
recently reported at the 2016 SVP meeting) in limestone paving slabs in a
sidewalk in Spain.

Fur seals and sea lions – also known as eared seals – are
numerically common in the North Pacific but arguably most diverse in the
southern hemisphere. Three of the five species of modern sea lions and all but
two species of fur seals live in the south. However, the fossil record of this
family (Otariidae) is almost entirely restricted to California and Japan, with
fossils in California extending back to the early Miocene (e.g. Eotaria).
One of the few anatomically informative fossils of an otariid from the southern
hemisphere is a nearly complete skull collected from Ohope Beach on the North
Island of New Zealand – studied first by JA Berry and subsequently named by
Australian pinnipedologist Judith King as Neophoca palatina. Neophoca is
the modern genus of Australian sea lion, and the New Zealand sea lion, Phocarctos
hookeri – does not have a pre-Holocene fossil record. This suggests that Neophoca
had a formerly wider distribution than the current species does, inhabiting
both Australia and New Zealand – or alternatively, inhabiting New Zealand prior
to dispersing to Australia. However, no pinniped specialists had even examined
the holotype skull since King named the species in 1983. Was the generic
allocation correct? We took the holotype skull on loan in 2013 while Morgan
visited New Zealand, and reconstructed it – it had shattered in the mail on
return from Australia, and we painstakingly glued many of the surviving bone
fragments together so that we could reassemble the skull for the first time in
30 years. Owing to cranial incompleteness of the holotype, earlier cladistic
analysis failed to resolve the position of Neophoca palatina within the
otariid tree. We instead used a principal components analysis to examine
relationships and found that cranial measurements successfully segregated all
southern otariids by genus. Neophoca palatina was confirmed using this
quantitative approach to be allied with extant Neophoca cinerea. This
suggests that Neophoca formerly had a wider range in temperature
tolerance and that New Zealand geography may have played an important role in
the evolution and dispersal of southern hemisphere otariids.

Xenorophid dolphins are a bizarre, highly derived and
short-lived monophyletic group of early odontocetes (toothed whales).
Xenorophids are known only from the Oligocene of South and North Carolina –
principally from the lower Oligocene Ashley Formation (SC) and the upper
Oligocene Chandler Bridge (SC) and Belgrade (NC) formations. In 2014, the new
genus and species Cotylocara macei was described, a xenorophid which
possessed a series of deeply excavated cranial sinuses and cranial asymmetry
indicating that it had the ability to produce high frequency sounds like modern
echolocating dolphins. But sound production is just one half of the equation –
were xenorophids capable of hearing high frequency sounds? Such an ability is
required for echolocation. In this study, Morgan Churchill and colleagues
report a new genus and species, Echovenator sandersi, which is a
somewhat smaller species of xenorophid. Echovenator lacks the derived
sinuses of Cotylocara, but the well-preserved earbones of Echovenator
permitted CT scanning of the cochleae – the organ of hearing embedded deep
within the periotic bone. Numerous features of the cochlea, including looser
cochlear coiling (tighter coiling = low frequency hearing, which characterizes
mysticetes), indicates that Echovenator – and likely all other
xenorophids – were capable of high-frequency hearing, and thus echolocation.
Because xenorophids are the earliest diverging odontocetes, this finding
indicates that echolocation evolved very early and likely represents a key
innovation driving the diversification of odontocetes. Lastly, the analysis
found that archaeocete whales possess adaptation for higher frequency sounds
than their ancestors, indicating that mysticetes and odontocetes diverged
towards high and very low frequency hearing at the Neoceti split, rather than
mysticetes inheriting low-frequency hearing from archaeocete ancestors with
odontocetes splitting off.

Barnacles are one of my favorite invertebrates, and along
the rocky shores of California,
they are practically everywhere; it's tough to find them here in South
Carolina because they generally require hard
substrate. Most barnacles - the weirdest of crustaceans - are adapted to
attaching to rocky substrate and occasionally to wood, making them pests for
boating/shipping - some are specialized towards "fouling" invertebrate
shells, and no doubt if you go to the lobster tank at the local grocery store
you'll probably find a barnacle or two on one of the crabs in there. Barnacles
either attach to substrate via attaching their carbonate skeleton directly to
the substrate by cement or protein (e.g. acorn barnacles), or by a soft tissue
stalk (e.g. gooseneck barnacles). Certain barnacles, however, have adapted
towards living on the shell of sea turtles and within the skin of whales
(chiefly baleen whales). Sea turtle shell provides less of a problem - it is
hard like rock, but because shell is coated in sheets of horn - keratin - which
exfoliate slowly during the turtle's life, turtle barnacles have to continually
pierce through layers in order to not be shed off. Whale barnacles often inhabit
much softer tissue, and many have evolved elongate "dagger" like
plates that pierce through the skin to maintain a hold as skin around the
barnacle periodically sloughs away. Owing to their unique and rare substrate,
both turtle and whale barnacles are as rare as hen's teeth in the rock record.
This paper reports a balaenid right whale skeleton from the Pliocene of Italy
with a number of turtle barnacles found preserved in association. These authors
suggest that rather than the skin, these turtle barnacles (Chelonibia
sp.) were attached to the callosities - large cornified white pads on the snout
and chin of right whales which house sensory hairs and host a wealth of
parasites (whale lice, and typically whale barnacles). Right whales are
relatively slow swimming with this unique hard tissue - permitting the jump
from turtle shell to a whale. More interestingly, Chelonibia is related
to whale barnacles and is an early diverging lineage within the group
Coronuloidea. This raises the possibility that marine turtles, which predate
pelagic whales by about 100 million years, may have served as an evolutionary
stepping stone - both intermediate in time and substrate quality - to whales.

The land to sea transition in the early evolution of whales
has been revealed by a series of well-preserved skeletons of pakiccetid,
ambulocetid, remingtonocetid, and protocetid archaeocetes largely from the
Tethys region and North Atlantic. These fossils show that early whales evolved
from terrestrial running ancestors, spent a fair amount of time in the water,
at first resembling wolves and eventually bizarre giant otter-like critters
with crocodile-like snouts (I lovingly refer to the remingtonocetids as the
“crocodile-otters”). The protocetids chronicle the move to marine settings and
the transformation of the hindlimb – later diverging protocetids lose fusion
between the sacral vertebrae, fusion between the innominate and the sacral
vertebrae, and ultimately the number of sacral vertebrae as well – prefacing
the decoupling of the hindlimb from the vertebral column and reduction in
hindlimbs in the basilosaurid archaeocetes. These aspects of the land to sea
transition were established entirely based upon the functional anatomy of the
skeleton as inferred from gross skeletal anatomy. However, in the last 15 years
or so, a number of studies reporting on bone microanatomy and histology as well
as stable isotopes have yielded further insights. Until this study, these sorts
of data have been considered in isolation but not together. This study reports
new microanatomical data (from CT and histology) and stable isotopes from
pakicetid and remingtonocetid whales as well as the curious deer-like cetacean
relative Indohyus. Stable isotopes indicate that pakicetids and Indohyus
likely inhabited freshwater settings (consistent with their preservation in
terrestrial deposits) and that remingtonocetids inhabited marine settings
(consistent with their marine sedimentological context, though terrestrial
mammals get swept out to sea and preserved in marine sediments on occasion).
Bone microanatomy suggested more variation, with some pakicetids being more
aquatic (Pakicetus – somewhat thicker cortex) and others more
terrestrial (Ichthyolestes – somewhat thinner cortex like a land
mammal), though cortical thickness in remingtonocetids was consistent with
aquatic habits. This study highlights the utility of both types of data in
inferring aquatic habits in cetaceans and other artiodactyls (e.g.
anthracotheres).

Beaked whales (ziphiidae)
are some of the more peculiar modern cetaceans - and cetaceans are already
quite strange as far as mammals go. Most ziphiids lack dentition other than
mandibular tusks, have enormous hyoid bones, and rather dense rostra. One
extant beaked whale, Blainville's beaked whale (Mesoplodon densirostris),
has the densest bone of any mammal (if not vertebrate) in its rostrum - and
apparently serves no function in diving or combat. The bottlenose whale, Hyperoodon,
bears paired fan-shaped crests formed from much spongier bone with a deep
furrow between for the melon. Fossil ziphiids expand this diversity and include
ziphiids with an inflated ridge down the top of the rostrum (Aporotus, Caviziphius,
Tusciziphius, Ziphirostrum), paired crests on the maxilla (Africanacetus,
Imerocetus) and perhaps the strangest of all - Globicetus hiberus,
with an enormous hemispherical tuberosity on the base of the rostrum. Earlier
histological studies showed that ziphiids independently grew all sorts of weird
cranial structures and used different growth pathways towards forming them. How
was the bizarre bony "soccer ball" formed on the rostrum of Globicetus?
These authors report the results of histological study of a referred skull of Globicetus
dredged off the continental shelf near Spain. Histology shows that the bone was
formed by continued periosteal accretion of the premaxillae long after the
whale reached physical and sexual maturity. The bone is extremely dense
(osteosclerotic) and laminar and lacks any evidence of remodeling. Curiously,
the orientation of bone growth seems to have rotated during growth of the
tuberosity. These authors conclude that the "internal antlers"
hypothesis proposed by Pavel Gol'din a few years ago (that weird sexually
dimorphic cranial structures are visible via echolocation and permit easy
recognition of species and sex to whales which spend much of their time at
depth) is the most likely explanation for rostral features in ziphiidae like
the rostral tuberosity in Globicetus.

Paleocetologists are crazy
about whale and dolphin earbones. They are loosely attached to the skull, and
typically fall out of the skull early on during decomposition. Externally, they
are weird shaped bones that are highly distinctive in shape and easily
identifiable, simultaneously permitting diagnosing new extinct species when
associated with more complete skeletal remains and permitting identification of
a particular species at a new locality when found in isolation. Internally, the
periotic, also known as the petrosal, houses the bony labyrinth – which
consists of the spiral-shaped cochlea and the semicircular canals. These
internal structures can only be seen by the naked eye if the specimen is broken
– and older studies destructively sampled these by serial grinding: literally
grinding away the bone micron by micron and taking hundreds of photographs or
camera lucida drawings. CT scanning is now routinely used in paleontology and
numerous studies have published on 3D models made from micro CT data of the
bony labyrinth of modern and extinct whales. I remember when I was an
undergraduate student, nearly everyone seemed to be doing this, but few
publications arose. Finally, many of these are being published, mostly by this
author (Eric Ekdale) and Rachel Racicot (below). This study presents new data
and analyses of modern and extinct baleen whale bony labyrinths. At a gross
level, this paper demonstrates that morphometric analysis of cochlear landmarks
is insufficient to clearly separate baleen whales and echolocating dolphins –
though when the semicircular canals are included, resolution becomes much
higher. This study supports an earlier paper by Ekdale and Racicot (2015) which
hypothesized that, based on cochlear morphology, basilosaurid archaeocetes
(which all Neoceti are thought to have evolved from) appeared to have
adaptations for low frequency hearing like modern mysticetes (but see Churchill
et al., above); the current study finds that Zygorhiza and the archaic
toothed “archaeomysticete” (from Charleston!) plot within the “morphospace” of
baleen whales. Other interesting findings: some cetotheriid baleen whales
including Herpetocetus and Metopocetus have extreme cochlear
coiling, and may have had some unknown super-low frequency hearing
specialization. Curiously, the middle Miocene allodelphinid dolphin Zarhinocetus
errabundus plots within baleen whale morphospace, near Metopocetus –
suggestive of low frequency hearing, or perhaps that certain extinct
odontocetes had a much wider range of hearing frequencies.

Fitzgerald, E.M.G. 2016. A late Oligocene waipatiid dolphin (Odontoceti: Waipatiidae) from Victoria, Australia. Memoirs of Museum Victoria 74:117-136.
In 1994 my former Ph.D. Adviser Dr. R.E. Fordyce found and
collected a particularly complete dolphin skull from a cliff exposure of the
upper Oligocene Otekaike Limestone near Duntroon, NZ. The specimen included a
cranium missing only the tip of the rostrum, both mandibles, earbones, and the
atlas vertebra. Oligocene dolphins are rare worldwide, and at the time, the
taxonomy of odontocetes was hopelessly confused; most heterodont cetaceans of
Oligocene age were assumed to be squalodontids, known from better presrved
Miocene deposits. Incomplete specimens and lazy taxonomy plagued the study of
dolphin evolution – so discovery of such a specimen offered a unique
opportunity to clarify the early evolution of dolphins. This new dolphin was
named Waipatia maerewhenua, and represented the Waipatiidae - one of now
several recognized groups of heterodont dolphins from the Oligocene. Other
waipatiids now include Otekaikea (NZ), Awamokoa (NZ), and
possibly Papahu (NZ) and Sulakocetus (Caucasus). A fragmentary
skeleton collected from the upper Oligocene Jan Juc Marl of Australia is
described in this new paper by Erich Fitzgerald and includes a partial
dentition and associated postcrania (the Jan Juc Marl also yielded the type
specimens of the mammalodontids Mammalodon colliveri and Janjucetus
hunderi). The teeth are quite small but clearly are separable into two
general categories: anterior conical teeth and posterior subtriangular cheek
teeth with accessory cusps. Some of the anterior teeth were even procumbent
tusk-like teeth, as seen in Waipatia maerewhenua and Otekaikea huata (named
for its spear-like tusks). The specimen preserves a nice forelimb – one of the
only known examples for an Oligocene odontocete. The combination of forelimb
features and the dentition identify this specimen as the first waipatiid from
Australia. More complete remains will eventually be found and further refine
the identification. Furthermore, some specimens from the Chandler Bridge
Formation of South Carolina (including CCNHM 107, our dolphin nicknamed
“whacky”, and a bunch of skulls I prepared this summer) likely represent
waipatiids from the North Atlantic. I expect future discoveries will illuminate
a worldwide geographic range for this group – but specimens from nearby
Australia are a critical first step.

Mammalodontid whales are the earliest recognized toothed
mysticetes, and include Mammalodon colliveri and Janjucetus hunderi
from the Jan Juc Marl of Australia – the same unit as the waipatiid reported by
Fitzgerald (see above). Mammalodontids are small (~3 m long) cetaceans with a
blunt rostrum, enormous eyes, and denticulate leaf-shaped cheek teeth; they are
the most primitive known mysticetes, and reflect early ecological
specializations within Mysticeti. Janjucetus is a seal-like apex
predator, and Mammalodon was likely a benthic suction feeder owing to
extreme tooth wear. Similar to the Fitzgerald article reporting the first
waipatiid from Australia, this paper reports the first mammalodontid from New
Zealand – Mammalodon hakataramea – based on a fragmentary skull
including the top of the braincase, a well-preserved tympanic bulla, and teeth
from the late Oligocene Kokoamu Greensand. The teeth are worn down to the
gumline and only roots remain; the bulla and skull are similar to M.
colliveri and differ only in a few minor details. The type specimen is,
however, the only published example of a NZ mammalodontid – and though a couple
other specimens needing reidentification exist (if you look hard enough in the
literature), I'm surprised by the rarity of mammalodontids in NZ. Mysticete
skeletons are not uncommon – eomysticetids and Mauicetus-like specimens
dominate Otago collections, and I've seen many more in the field – but only a
few precious scraps of mammalodontids. Then again, toothless mysticetes and
kekenodontids are not yet reported from the Jan Juc Marl (though the purported
kekenodontid “Squalodon” gambierensis is known from the early
Oligocene Gambier Limestone of Australia). This discovery extends the range of
mammalodontids to New Zealand – perhaps unsurprisingly, the Oligocene whale
assemblages are beginning to resemble one another more and more.

Baleen is the organ which baleen whales use during filter
feeding to strain tiny prey items from the water column – and contrary to
popular belief, relatively little of it consists of microscopic plankton – most
prey of baleen whales are amphipods, krill, and even schooling fish. Baleen is
not part of the skeleton or dentition – though in the mouth, it is a structure
unrelated to teeth, and is instead made out of keratin – the same protein that
skin, hair, claws/hooves, and horns are composed of within most vertebrates.
Like horns, keratin grows continuously as it is abraded and worn down by the
tongue. Baleen is lightly mineralized, but because it is a soft tissue, it is
rarely found in the fossil record. It has been found at a few localities in the
eastern Pacific: Purisima Formation (California), Monterey Formation
(California), Empire Formation (Oregon), and most importantly, in numerous
skulls and skeletons from the Pisco Formation of Peru. Because soft tissue
preservation is unexpected relative to skeletal preservation, what processes
permit baleen preservation? This new study reports the first study of baleen
preservation using a combination of microscopy and X-ray diffraction. This
analysis indicates that early diagenetic formation of dolomite – a type of
carbonate mineral similar to limestone – rapidly formed around the carcass and
preserved a natural cast around the baleen plates. Preservation is good enough
to show individual baleen tubules. Some microstructure of the baleen was
presrved by phosphatization, either during or after precipitation of dolomitic
cement around the baleen and skeleton. Furthermore, this study indicates that
baleen preservation was mediated by sinking of the carcasses into soft, soupy
sediment as hypothesized for exceptional preservation of Jurassic ichthyosaurs
in the Posidonia Shale of Germany – directly contradicting diatomite deposition
rates published for Pisco whales by young earth creationists, founded with
intent to disprove radiometric dating and prove a young, biblical age of the
earth.

Sometime in the 1840s a curious skull was dug out of a bank
near Middleton Place plantation right here in Charleston, SC, only a few miles
west of my apartment in West Ashley. The skull was studied initially by various
naturalists including Louis Agassiz, eventually named Agorophius pygmaeus (though
other names have been applied to the same skull, e.g. Phocodon holmesii)
but disappeared sometime in the late 19th century. Later researchers determined
that, based on the beautifully illustrated published plates of the specimen,
that it represents an echolocating toothed whale (Odontoceti), albeit
interpreted as an archaeocete by earlier authorities; based on the location and
the morphology, the specimen likely originated from the lower Oligocene Ashley
Formation. The whereabouts of the skull are unknown, and Dave Bohaska thinks it
may be rediscovered if the historical trash heaps at the estate Agassiz was
staying were ever excavated by archaeologists. In 1980, my former Ph.D. adviser
R.E. Fordyce was working on his postdoc at the Smithsonian, and on a trip to
Harvard he found an isolated, mislabeled tooth that matched the single tooth of
the holotype (again, thanks to the highly detailed illustrations). This,
unfortunately, is the only surviving part of the holotype. New specimens
reported in this study - certainly one of the publications I was happiest to
see this entire year - alleviate this problem. Godfrey et al. refer two new
skulls to Agorophius pygmaeus, both including well-preserved braincases
and one with a nice periotic (earbone). The morphology more or less conforms to
the published figures, but gives us crucial new reference specimens to base the
genus and species on. A similar problematic example is Zygorhiza kochii -
the holotype is garbage, but we colloquially use the most complete skeleton and
cranium (USNM 11962) as the de facto reference specimen rather than the type. we
can now do this for Agorophius, the concept of which has been based on nothing
more than illustrations and a tooth for over a century. The new material
permits inclusion of Agorophius pygmaeus within a cladistic analysis, and in
the analysis reported by Godfrey et al., Agorophius plots out low on the
odontocete tree, between the Xenorophidae and more derived odontocetes like the
"squalodontids" and Waipatia. We have more material of Agorophius
pygmaeus here in CCNHM collections... so stay tuned!

The last decade has
witnessed a revolution in the study of "cetotheres" - primitive
baleen whales within the crown group that evidently do not belong to any modern
family (but see Marx and Fordyce, below). In the 19th and 20th century, a
number of extinct chaeomysticetes were named and lumped into the wastebasket
"Cetotheriidae" - including species now known to be eomysticetids
(e.g. Tokarahia lophocephalus, Micromysticetus), and by the 1990s
it was widely acknowledged that "cetotheriids" were either
paraphyletic or polyphyletic, but few researchers had anything constructive to
say about it. In 2006, a new study on the peculiar dwarf mysticete Piscobalaena
emanating from Virginie Bouetel's Ph.D. thesis included a cladistic analysis of
mysticetes, and found that a number of "cetotheres" formed a clade
with Cetotherium rathkii - including Herpetocetus, Nannocetus,
Piscobalaena, Metopocetus, and Mixocetus. Recognition of
this clade revitalized the study of "cetotheres", and given the
inclusion of the type species of the horribly overbloated Cetotherium,
the family Cetotheriidae was redefined. A number of additional studies by
Ukrainian cetologist Pavel Gol'din and Russian paleontologist Konstantin
Tarasenko have reported new genera and species as well as reevaluated old 19th
and early 20th century taxa. This new study by Gol'din and Startsev clarifies
the taxonomy and provides new descriptions for many problematic and poorly
described late Miocene taxa from the northeastern margin of the Black Sea
(Crimean peninsula and Caucasus), including the new genus Mithridatocetus,
which includes the new species M. eichwaldi and Mithridatocetus adygeicus
(originally placed in Kurdalogonus). "Cetotherium" mayeri
is recombined as Mithridatocetus sp., and Kurdalogonus is left
with only the type species, K. michedlizei. Additional species are
designated as nomina dubia (Cetotherium priscum and Cetotherium
incertum) and others simply require additional material and study to be
reevaluated ("Cetotherium" klinderi, "Cetotherium"
pusillum, "Cetotherium" maicopicum, Eucetotherium
helmersenii). A fascinating result of this research is that most of
these taxa - true Cetotherium, Kurdalogonus, Mithridatocetus,
Eucetotherium, and Brandtocetus - form a clade restricted to
Paratethys (a redefined Cetotheriinae), suggesting a short-lived endemic
radiation of cetotheriids within the Paratethys. These authors speculate that
the endemic mysticete fauna likely went extinct owing to the warming associated
with the Messinian Salinity Crisis (for the uninitiated, a period of time where
the Mediterranean almost completely evaporated).

Govender, R., M. Bisconti, and A. Chinsamy. 2016. A late Miocene-early Pliocene baleen whale assemblage from Langebaanweg, west coast of South Africa (Mammalia, Cetacea, Mysticeti). Alcheringa 40:4:542-555.
Few studies of marine
mammals document assemblages from a faunal perspective – many palentologists
tend to focus on a subset of taxa, which has an unfortunate side effect –
marine mammal assemblages are rarely published in their entirety. I discussed
this a bit in my 2013 Purisima Fm. marine mammal monograph. The world's best
known marine mammal fossil assemblage – the Pungo River Limestone and Yorktown
Formation of the Lee Creek Mine, North Carolina – has been extensively
published on, and followup papers to a series of monographic works in the Lee
Creek Mine edited volumes are supplementing published material with newly
discovered specimens and revising identifications. Another similar assemblage
of equivalent early Pliocene age like the Yorktown Fm. - and, also from a strip
mine – is the assemblage from Langebaanweg in South Africa. This assemblage is
well known for fossils of the lobodontine seal Homiphoca capensis –
however, few, if any, cetaceans have been reported from the assemblage up until
now. This study reports baleen whale earbones from the mine, mostly
identifiable as whales similar to the late Miocene Italian rorqual Plesiobalaenoptera
quarantellii. Interestingly, perhaps three species of archaic rorqual may
be present. The earbones possess some features similar to gray whales,
including a fenestra rotunda that is teardrop shaped and open dorsally. More
complete material is obviously needed in order to name any species.

Kekenodontids are poorly known archaic cetaceans from the
Oligocene – founded upon the fragmentary basilosaurid-like Kekenodon onamata
from the upper Oligocene Kokoamu Greensand of the South Island of New
Zealand. Other possible kekenodontids include Phococetus vasconum from
the Oligo-Miocene of France and “Squalodon” gambierensis from the
Oligocene Gambier Limestone of Australia. These latter taxa are known only from
isolated teeth – though a recent SVP presentation by my former labmate Josh
Corrie reports new material of “S.” gambierensis from New Zealand
which firms up a link with Kekenodon. Most of the research on
kekenodontids is unpublished (Corrie, in prep) but indicates these critters are
basilosaurid-like with some features of Neoceti, and likely to be late
surviving archaeocetes rather than early archaeocete-like toothed mysticetes.
This new study reports a very archaic periotic from the upper Oligocene El Cien
Formation of Baja California – equivalent in age with the Yaquina and Pysht
formations of the Pacific Northwest, and already known to yield aetiocetids and
eomysticetid-like baleen whales. This new periotic has a rather high superior
process – a ridge that articulates with the squamosal, characteristic of
basilosaurid archaeocetes. Several features of the pars cochlearis are shared
only with Kekenodon onamata – which is known from a pretty weird-looking
periotic (in addition to a bulla, frontal, quite a lot of the dentition, and an
atlas). I'm not necessarily convinced just yet – and more complete material
from Baja is needed. Furthermore, Kekenodon onamata, and other new
kekenodontids, await description from New Zealand (get on it Josh Corrie!).
Here is yet another paper on
fossil beaked whales this year (see also Bianucci et al., Dumont et al., and
Mijan et al.). Dredged ziphiids have been extensively reported by now – and one
of the first major descriptions of a dredged assemblage was published by
Bianucci et al. (2007) on a diverse assemblage of Miocene or Pliocene ziphiid
crania from the continental shelf off of South Africa. One ziphiid they named, Africanacetus
ceratopsis, has a relatively elongate, narrow rostrum somewhat similar to Mesoplodon
but has a pair of conical “horn”-like tuberosities on the maxilla. Africanacetus
is most closely related to the bottlenose whale Hyperoodon, which
similarly sports paired maxillary crests. This study reports a new species of Africanacetus
collected by an ROV from a slope of the Sao Paulo Ridge off the shore of
Brazil. It's represented by a well-preserved skull coated in manganese oxide –
a common mode of preservation for deep marine cetacean specimens (in contrast,
the South African specimens were coated in phosphate nodules, much more similar
to preservational modes in continental deposits of shallow marine sediments.
Additionally, the skull was found in a field littered with manganese nodules.
Isotopic analysis of the manganese indicates the skull sat exposed on the
seafloor for at least the last five million years, with manganese oxide
deposition commencing about 5 Ma. A maximum age of middle Miocene – the period
of time when uplift of the Sao Paulo Ridge terminated. This new species, Africanacetus
gracilis, differs in minor ways from Africanacetus ceratopsis as
well as being more gracile in skull proportions (hence the species name). Like
many other extinct and nearly all modern ziphiids, Africanacetus lacks an upper
dentition and was likely a suction feeder specializing on squid. Discovery of Africanacetus
this close to the equator, and relatively close to Antarctica (Gol'din and
Vishnyakova, 2013) suggests that this taxon had a relatively widespread
distribution during the Miocene.

In the 1930s the cetacean assemblage from the lowermost Miocene
Jewett Sand near Bakersfield was reported, and one of the better specimens
consisted of a partial braincase with earbones and postcrania named Allodelphis
pratti. In the past 20 years, a number of new specimens have been
collected, which have allowed naming of the new species Allodelphis
woodburnei, and reassignment of the problematic taxon “Squalodon” errabundus
from the Sharktooth Hill Bonebed to the new genus Zarhinocetus.
Allodelphinids are smallish, long-snouted dolphins with a number of features allying
them with platanistids (Ganges river dolphin and extinct relatives). This new
study critically expands the fossil record of allodelphinids, and reports many
new specimens and taxa. More detailed descriptions of Allodelphis spp.
and Zarhinocetus errabundus are provided, and the new species Zarhinocetus
donnamatsonae is described from the lower Miocene Astoria Formation of
western Washington. A beautifully preserved skull with articulated mandibles
from the lower Miocene Nye Mudstone of Oregon is named as Goedertius oregonensis
after “amateur” paleontologists Jim and Gail Goedert who collected the
specimen. A new allodelphinid, Ninjadelphis ujiharai, is also
named from a partial skull with earbones and vertebrae the lower Miocene Awa
Group of Japan. Allodelphinids share similarly long rostra, bizarrely curved
zygomatic processes of the squamosal, elongated and enlarged cervical
vertebrae, and a secondarily elongate humerus. Allodelphinids represent an
Oligocene-middle Miocene diversification of longirostrine platanistoid
odontocetes that never left the North Pacific and went extinct sometime after
the middle Miocene.

This summary is going to be brief owing to the fact that the
original article is entirely in Japanese (which I cannot read). This study
reports a rare example of an articulated odontocete forelimb, and includes the
distal radius, ulna, carpals, metacarpals, and phalanges. Details of carpal
articulations are described in detail – and carpals rarely fossilize, likely
owing to disarticulation of the flipper during prolonged floating of carcasses.
Unfortunately, functional evaluation within a phylogenetic context is not
possible since the skull is missing.

This problematic attempts to resurrect the largely discarded
idea of pinniped diphyly – the hypothesis that modern pinnipeds evolved from
two completely different groups of arctoid carnivores: eared seals and walruses
(Otarioidea) from bears, and earless seals (Phocidae) from mustelids (weasels
and otters). The idea was first posited in the 1950s and adopted by most
pinniped paleontologists in the 1960s, and remained more or less unchallenged
until a series of papers were published by Andre Wyss and Annalisa Berta on
pinniped osteology and paleontology. These papers included the first
computer-run cladistic analyses of morphological character data, and proposed
abundant evidence for pinniped monophyly. Shortly thereafter, molecular
analyses unequivocally supported pinniped monophyly as well – and though this
new paper cites a few oddball examples of
early karyological and molecular studies supposedly not supporting
pinniped monophyly, literally dozens upon dozens of molecular analyses
published since the early 1990s on pinniped and carnivore relationships have
supported pinniped monophyly. This study provides a reinterpretation of several
“important characters”, some of the anatomical interpretations being quite
problematic. For example, many cranial characters supporting monophyly are
argued ad nauseum with emhasis on how different each pinniped family is from
one another; shortening of the humerus and femur is argued as being
uninformative because it also characterizes other marine tetrapod groups.
Shortening and anteroposterior flattening of the femur is argued away because this is claimed to characterizes
penguins – however, all of the penguin femora I've seen are perhaps shortened,
but have a cylindrical shaft. According to these authors, many features cannot
be used in cladistic analysis because they are convergent in phocids and
“otarioids” - however, how are we supposed to know? These authors poke holes,
some of which are valid and suggest some refining of character definitions, but
ultimately we're supposed to just take their word for it. If the character
evidence is truly convergent, a cladistic analysis incorporating as much data
as prior analyses supporting monophyly should in theory produce a diphyletic
result. However, the cladistic analysis they report includes a paltry seven
taxa coded for 12 characters – a small fraction of the character data published
23 years ago by Berta and Wyss (1994). With such a small matrix, it begs two
questions: why were the authors unable to find more character evidence
supporting diphyly? And is the matrix small because the authors cherry picked
available characters in order to produce the result they wanted? Neither option
is desirable.

Beaked whales are perhaps one of the most surprisingly
diverse groups of modern cetaceans. Many new species have been discovered,
proposed, and named within my lifetime despite never being seen alive – rare
and unnamed species are often discovered entirely based on strandings. Earlier
this year, a dwarf species (as yet unnamed) of the giant beaked whale genus Berardius
was reported from Alaska based on stranded specimens. The fossil record of
baked whales is now extensive, yet most of the record consists of reworked
specimens dredged from seafloor deposits of unknown age, and few records of
extant genera in the fossil record are known. Molecular divergence dating
offers a method to estimate the timing of phylogenetic divergences – yet
changes in the rate of molecular evolution mean that parts of the tree need to
be calibrated with fossil occurrences with good dates. Because few fossils
nested within the part of the tree that includes modern species exist, few
calibrations exist for Ziphiidae. This study reports a new species of the
modern genus Mesoplodon from the lower Pliocene, Mesoplodon posti,
named after Dutch paleontologist Klaas Post. A series of skulls are referred to
this new species, all of which were collected by unknown collectors in the late
19th century. Fortunately, associated matrix indicates that most of
the specimens were collected from the lower Pliocene Kattendijk Formation
(3.9-4.9 Ma). This is the first time good age control is available for fossil Mesoplodon,
indicating that diversification of Mesoplodon had begun by at least the early
Pliocene.

Modern sperm whales (Physeteroidea) are highly specialized
deep divers that feed almost exclusively on squid. Some extinct physeteroids
exhibited similar feeding adaptations, but an increasingly well-documented
group of early sperm whales sport enormous teeth with enamel (lost in extant
physeteroids) and robust rostra and mandibles. Zygophyseter, Brygmophyseter,
and Acrophyseter were discovered, and eventually the gigantic Livyatan
melvillei - similar in size to modern Physeter (giant sperm
whale), but with upper and lower teeth the size of 2 liter beverage bottles and
a robust skull the size of a small car - was discovered in the Peruvian desert.
Even more recently, Albicetus oxymycterus from the middle Miocene
of was redescribed, just a bit smaller than Livyatan. This study is a
follow-up to the original paper on Livyatan in Nature, and includes a
more detailed anatomical description, as well as a similar description of Acrophyseter
deinodon - also from the Pisco Formation. Earbones of Acrophyseter are
described for the first time. The authors also report a new species, Acrophyseter
robustus, a species somewhat older and with a more robust skull than Acrophyseter
deinodon. A third possible species is recognized but not named (Acrophyseter
sp.). All of these large-toothed sperm whales are killer whale-like and were
the apex predators in the Miocene - with competition for prey only from the
giant shark Carcharocles megalodon. Cladistic analysis interestingly
supports recognition of a clade including Acrophyseter spp., Brygmophyseter,
and Zygophyseter, as one of the earliest diverging physeteroid clades; Livyatan
is positioned one node crownward, as sister to the Physeteridae + Kogiidae
clade. Livyatan and Acrophyseter likely coexisted, suggesting
some degree of niche partitioning - for example, that each preyed upon marine
vertebrates appropriate for their size.

Modern walruses are one of the largest pinnipeds and are
also simultaneously the most charismatic owing to their bizarre feeding
apparatus, including a formidable pair of tusks formed from enlarged canines.
The modern walrus is an effective suction feeder and has a voracious appetite
for bivalve mollusks (clams) - and does not even use its teeth; it simply sucks
the flesh right from the clam shell. Other extinct walruses (Valenictus,
Ontocetus, Aivukus, Gomphotaria) are similarly interpreted
as being molluskivirous, however this is a recent event in walrus evolution -
most walruses were remarkably sea lion-like for the majority of their evolution
(~5-17 Ma). One middle Miocene walrus in particular, Pelagiarctos thomasi,
was initially interpreted to be a "killer walrus" that fed on marine
mammals and perhaps seabirds. In a 2013 paper, Morgan Churchill and I found
that on morphological and relative abundance evidence, Pelagiarctos was
unlikely to be anything but a generalist fish eater like most modern seals and
sea lions and found no anatomical adaptations for feeding on large prey. While
still in New Zealand, labmate Carolina Loch approached me about sampling Pelagiarctos
for an SEM study of enamel ultrastructure as another way to test the
"killer walrus" hypothesis. We sampled the enamel of a new tooth of P.
thomasi collected by colleague J.P. Cavigelli, as well as the New Zealand
sea lion and fur seal for comparison - both extant fish-eating generalists.
Hunter-Schreger bands are zigzag-like convoluted bands in enamel that prevent
cracks from propagating; mammals that bite hard prey tend to have this
developed more strongly. Instead, there was not really much of a difference
between any of the three pinnipeds - leading us to propose that Pelagiarctos
lacks obvious ultrastructural adaptations in its enamel for feeding on
large fish, and is no more specialized than extant otariids.

Marx, F.G. and N. Kohno. 2016. A new Miocene baleen whale from the Peruvian desert. Royal Society Open Science 3:160542.
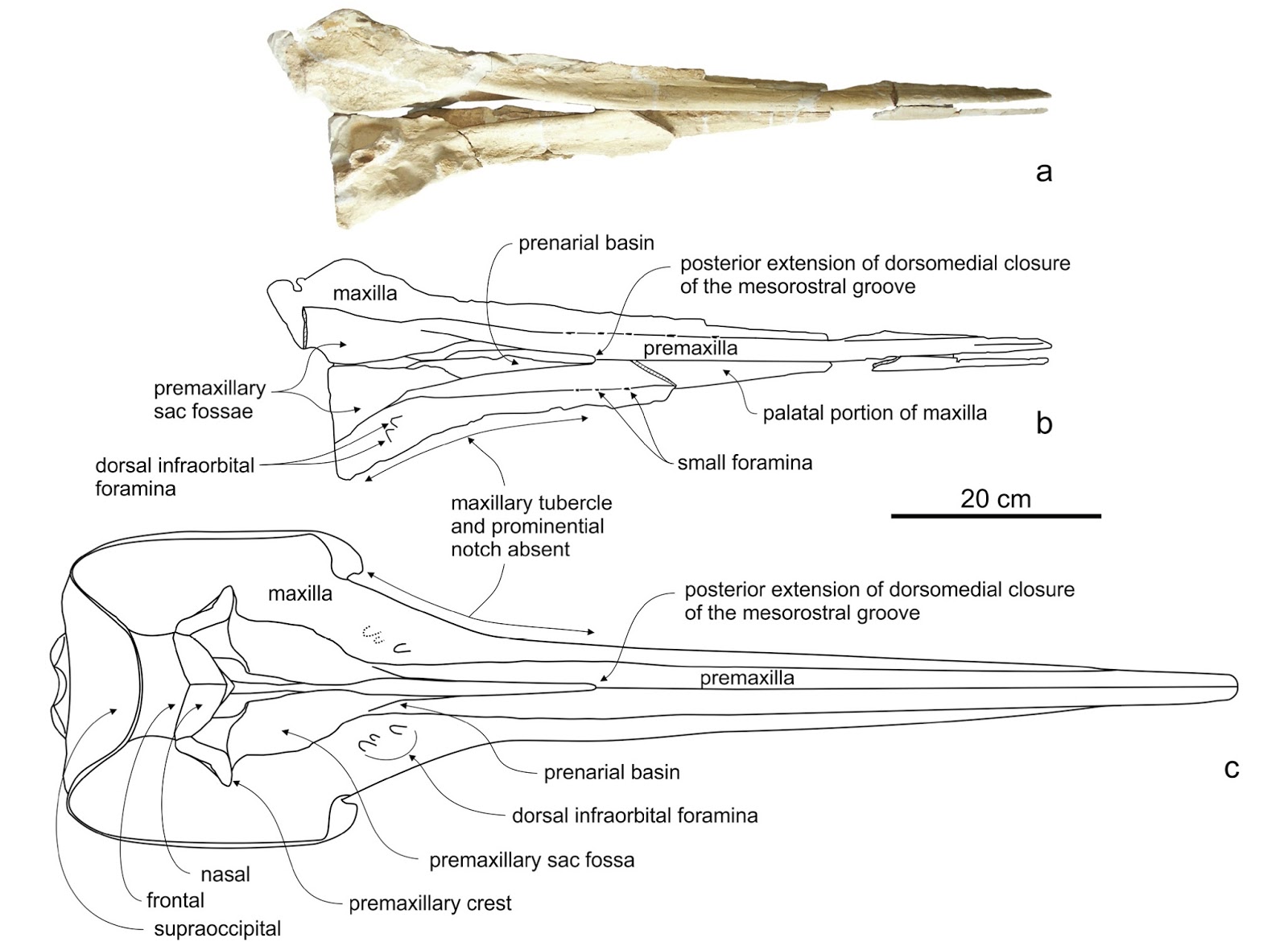
The fossil record of balaenopterids has been plagued by a problematic history, including specimens which have been assigned too damn many names in the literature (e.g. Pliocene balaenopterids from Italy) and taxa based on too damn many specimens, often chimaeric in nature (e.g. Pliocene balaenopterids from Belgium). Many are poorly figured and present in museums on different continents - meaning that paleontologists with local focus, or young paleontologists, are unable to really publish anything meaningful on new balaenopterid material if 1) unable to visit these collections and 2) unable to use the available literature. As a result, there is an embarrasingly large volume of undescribed balaenopterid skulls and skeletons across the globe, chiefly including California, Oregon, Florida, North Carolina, Japan, Peru, New Zealand, Australia, Italy, and the North Sea region. A number of recent studies have begun to erode away at this problem, both by fixing ancient taxonomic problems and redescribing old material and by describing new, beautifully preserved specimens. This new study reports the new genus and species of Incakujira anillodefuego - the genus name referring to the Peruvian origin of the specimens and their current disposition in Japan (Kujira is "whale" in Japanese) and the species name referring to the the Pacific ring of fire, which both Japan and Peru sit upon. Incakujira is based upon two absolutely gorgeous specimens from the upper Miocene Pisco Formation of Peru, and is characterized by having a relatively narrow rostrum, slender ascending maxillae, elongate nasals, and lacking a squamosal crease (a synapomorphy of Balaenoptera). Many aspects of the skull are similar to the modern humpback whale, and indeed the cladistic analysis supports placement of this species as sister to extant Megaptera. This analysis used a molecular partition, which places Megaptera within Balaenoptera, making the latter paraphyletic. Incakujira is inferred to be a lunge feeder like modern balaenopterids, though it possesses a twisted postglenoid process which may suggest an ability to skim feed. The holotype of Incakujira preserves baleen, and spacing of baleen is similar to Balaenoptera acutorostrata (minke whales).
The fossil record of balaenopterids has been plagued by a problematic history, including specimens which have been assigned too damn many names in the literature (e.g. Pliocene balaenopterids from Italy) and taxa based on too damn many specimens, often chimaeric in nature (e.g. Pliocene balaenopterids from Belgium). Many are poorly figured and present in museums on different continents - meaning that paleontologists with local focus, or young paleontologists, are unable to really publish anything meaningful on new balaenopterid material if 1) unable to visit these collections and 2) unable to use the available literature. As a result, there is an embarrasingly large volume of undescribed balaenopterid skulls and skeletons across the globe, chiefly including California, Oregon, Florida, North Carolina, Japan, Peru, New Zealand, Australia, Italy, and the North Sea region. A number of recent studies have begun to erode away at this problem, both by fixing ancient taxonomic problems and redescribing old material and by describing new, beautifully preserved specimens. This new study reports the new genus and species of Incakujira anillodefuego - the genus name referring to the Peruvian origin of the specimens and their current disposition in Japan (Kujira is "whale" in Japanese) and the species name referring to the the Pacific ring of fire, which both Japan and Peru sit upon. Incakujira is based upon two absolutely gorgeous specimens from the upper Miocene Pisco Formation of Peru, and is characterized by having a relatively narrow rostrum, slender ascending maxillae, elongate nasals, and lacking a squamosal crease (a synapomorphy of Balaenoptera). Many aspects of the skull are similar to the modern humpback whale, and indeed the cladistic analysis supports placement of this species as sister to extant Megaptera. This analysis used a molecular partition, which places Megaptera within Balaenoptera, making the latter paraphyletic. Incakujira is inferred to be a lunge feeder like modern balaenopterids, though it possesses a twisted postglenoid process which may suggest an ability to skim feed. The holotype of Incakujira preserves baleen, and spacing of baleen is similar to Balaenoptera acutorostrata (minke whales).

This new book is a welcome
text on the evolution and fossil record of whales and dolphins. An enormous
volume of research has been published in recent years, with perhaps half of the
studies on fossil cetaceans published within my lifetime – and the science is
progressing at such a rapid pace that new review articles are being published
almost once a year (whether we're asking for them or not). While I've been
privately critical of the need for such review articles, this new text is not
indulgently long (just over 300 pages) and includes introductory chapters, a
pertinent chapter on modern cetaceans, methods used in cetacean paleontology,
chapters on functional morphology, phylogenetics, paleoecology, biogeography, and
cetacean macroevolution. Summaries of each cetacean family and other clades are
succinct, comprehensive, highly informative, and supplemented with dozens of
new illustrations all of consistent style (which greatly pleases my anal
retentive tendencies). The book also includes a number of gorgeous color
illustrations by Carl Buell. I've not read the book in full, but am seriously
impressed with the depth of knowledge, comprehension, and level of detail. This
book is a must-have for paleocetologists new and established.

Baleen whale fossils are
some of the more commonly preserved marine mammals, and their large size
ensures that they are some of the first fossils to be found in an area with
fossiliferous Cenozoic outcrops. Indeed, legendary paleontologist Edward
Drinker Cope published extensively on many early baleen whale discoveries from
the Miocene Chesapeake Group of Maryland and Virginia. One of these is a
peculiar skull which he named Metopocetus durinasus – which was found ex
situ on a riverbank. Metopocetus has nasals fused at the midline, and a
plug-like posterior process of the periotic like Herpetocetus. Metopocetus
is now known to be a cetotheriid whale closely allied with Herpetocetus,
Piscobalaena and Cetotherium – but its age is completely unknown.
It possibly originated from the mid Miocene Calvert Formation, but similar
mysticetes of equivalent evolutionary grade occur within the younger upper
Miocene St Marys Formation. This study reports a new species of Metopocetus,
M. hunteri, from the upper Miocene of the Netherlands. It differs in a
few minor features of the periotic, and also includes a tympanic bulla and more
of the basicranium than M. durinasus.Both species are characterized by a
deep fossa on the paroccipital process in the basicranium, which they interpret
as a fossa for the stylohyal. Lastly, the age of this new species is well known
– 8.8-7.6 Ma, or Tortonian equivalent. This suggests that Metopocetus
durinasus likely originated from the younger St. Mary's Formation.

Approximately one year ago, Marx et al. named the new
toothed mysticete (Aetiocetidae) Fucaia
buelli, and proposed that large gums preceded baleen. Other toothed mysticetes
in the family Aetiocetidae have a series of minute palatal foramina which are
homologous with the larger and more extensive foramina present in modern
mysticetes. This was originally published by Tom Demere and others in 2008
about the remarkably well-preserved holotype skull of Aetiocetus weltoni from
the Oligocene Yaquina Formation of Oregon – showing that some toothed
mysticetes exhibited teeth and baleen. This new study reports a nice cranium
(collected by Jim Goedert) of an as-yet unnamed aetiocetid that is
morphologically intermediate between Fucaia
and Aetiocetus from the Pysht
Formation of Washington. This specimen has a peculiar style of tooth wear – the
lowermost part of the enamel crowns are pristine and unworn, whereas the tips
of the teeth are highly worn – indicating that the base of the crowns were
embedded in thickened gums (normally, the base of the enamel crown marks the
gumline). Furthermore, the wear is manifested on the mesial and distal edges of
the teeth – suggestive of sediment moving between the teeth. These authors
rightly interpret this to be evidence of benthic suction feeding. We made a
similar case for benthic suction feeding in the bizarre porpoise Semirostrum ceruttii (Racicot et al.,
2014). They further suggest that rather than possessing baleen, all aetiocetids
possessed thickened gums instead as an adaptation for suction feeding – and
that palatal vascularization originated first to supply blood to enlarged gums,
and the gum tissue was later co-opted for filter-feeding baleen-like structures
(modern baleen is derived from gingival tissue after all). This would indicate
that eomysticetids – the group I studied for my Ph.D. – where the first
cetaceans to be obligate filter feeders. They point out that having teeth and baleen
simultaneously presents problems as teeth would damage/interfere with baleen –
though I’m not really convinced. They make a good case for suction-based
feeding with enlarged gums based on behavior in this one unnamed taxon.
However, it is still possible that baleen was present in Aetiocetus proper, and given that analogous tooth wear is not
present in any Aetiocetus spp., a
different method to look for evidence of enlarged gums would be necessary.

It's been a pretty damn good
year for beaked whale paleontology (see Bianucci et al., Dumont et al.,
Ichishima et al., Rammasamy) - following several years of some rather
spectacular beaked whale fossil discoveries. One of the most unexpected sources
of beaked whale fossils have been offshore dredging, which have recovered all
sorts of bizarre critters (e.g. Globicetus; Dumont et al., above). Not
all discoveries reveal spectacular new skeletons representing unknown genera,
however – but some seemingly minor discoveries often serendipitously help solve
century-old problems and questions. This study reports a new species of the
archaic ziphiid Beneziphius. The new species, B. cetariensis, was
dredged from offshore Spain – the same area as Globicetus, Choneziphius
leidyi, and Tusciziphius atlanticus. Beneziphius has a
generalized Ziphius-like skull in many respects yet belongs to the
extinct “Messapicetus clade” (Bianucci et al., above). Like other dredged
specimens, the age of the holotype of B. cetariensis is not precisely
known. The genus Beneziphius was originally named based on a skull from
Belgium – but because it was collected without locality information in the 19th
century, its age is unknown as well. The desire for clarified age control for
this genus led the authors to sample matrix hidden inside the cranial cavity of
the 19th century holotype of Beneziphius brevirostris. Matrix
yielded Serravallian-aged (late middle Miocene) dinoflagellate cysts and
acritarchs (microfossils useful for biostratigraphy). This study further
remarks upon the faunal similarity of the Iberian dredged assemblage and the North
Sea – both include Beneziphius, Caviziphius, Choneziphius,
and Ziphirostrum; however, the Iberian assemblage includes Tuscizphius,
formerly reported from Italy
and South Carolina – but not the North
Sea.

Murakami, M. 2016. A new
extinct inioid (Cetacea, Odontoceti) from the upper Miocene Senhata Formation, Chiba,
central Japan:
the first record of Inioidea from the North Pacific Ocean.
Paleontological Research 20:3:207-225.
Modern river dolphins
include four different species, formerly grouped into the Platanistoidea – and
subsequently regarded as being a paraphyletic or polyphyletic assemblage. The
Platanistoidea is now redefined as Platanista (Ganges
river dolphin) and extinct relatives. The two river dolphins unique to South
America – Inia, the Amazon river
dolphin, and Pontoporia, the Franciscana or La
Plata river dolphin – likely are closely related and
frequently grouped into the Inioidea. True inioids are generally unique to the Americas
but some far-flung (but fragmentary) examples have been reported from western
Europe and the Mediterranean. This study reports a
fragmentary inoid dolphin skull from the Senhata Formation of Japan – named Awadelphis
hirayamai. This species is characterized by a laterally overhanging premaxillary
eminence, unique amongst inioids, a short zygomatic process, a posteriorly
enlarged nuchal crest, and other features of the facial region. This new taxon
is phenetically similar to pontoporiids yet Inia plots out within the
Pontoporiidae in the cladistic analysis – and thus Murakami refers the new
genus only to the Inioidea rather than a more exclusive clade within the group.
Awadelphis extends the range of inioids to the western North Pacific –
though the lipotid dolphin Parapontoporia has been preliminarily
reported, formerly known only from California
and Baja California. Curiously,
despite this other record, no comparisons were made with the lipotid Parapontoporia
– which was originally confused for a pontoporiid dolphin owing to some
convergent features.

Nance, J.R., M. Kricun, and S.J. Godfrey.
2016. Ankylosis and oosteonecrosis in the pectoral limb of a baleen whale
(Cetacea, Mysticeti) from the Miocene Calvert Formation of Calvert Cliffs, Maryland,
U.S.A. Marine Mammal Science. Online early.
Obligate marine mammals like
whales and dolphins and sea cows do not have to support their weight on land
(see Ando and Fujiwara, above). Because of this, certain injuries may be more
prevalent in marine mammals – not because they are more susceptible to injury,
but because they likely have a higher chance of surviving an injury. Skeletons
of modern marine mammals frequently have broken bones – including vertebrae
that are fused together. This new study reports a fossil baleen whale forelimb
from the middle Miocene Calvert Formation of Maryland. This specimen includes a
humerus, radius, and ulna – all fused together at the elbow. The radius is
quite enormous, and when examined via CT scanning, is osteonecrotic on the
interior along the entire length; smaller osteonecrotic zones are present in
the ulna and humerus. Unfortunately, the forelimb was found isolated, so the
extent of the injuries in the skeleton is unknown. What could have caused
osteonecrosis (bone death) and fusion of the elbow in this extinct baleen
whale? The authors interpret fusion of the elbow joint as a result of
immobilization of the elbow caused by osteonecrosis of the forelimb bones.
Osteonecrosis itself was likely specifically dysbaric osteonecrosis – caused by
decompression syndrome, otherwise known as “the bends”, as already documented
in other middle Miocene baleen whales.

Park, T., E.M.G. Fitzgerald, and A.R. Evans. 2016.
Ultrasonic hearing and echolocation in the earliest toothed whales. Biology
Letters 12:20160060
http://rsbl.royalsocietypublishing.org/content/12/4/20160060
Modern baleen whales (Mysticeti) and toothed whales (Odontoceti)
differ from one another in many regards, and aside from teeth v. baleen the
most important features differentiating the two are related to hearing. As
described above (Churchill et al.), echolocating odontocetes are adapted to
producing and hearing high-frequency sounds, whereas baleen whales are adapted
toward hearing low-frequency sounds and are incapable of echolocating. Some
studies using CT data have found evidence of high frequency hearing in Miocene
and Pliocene odontocetes - but how early did it evolve? This new study reports
CT data from the cochlea of an isolated periotic of a xenorophid dolphin -
xenorophids are some of the most common odontocetes in the Oligocene deposits
here in the Charleston embayment, and are the earliest diverging clade of
odontocetes. The bizarre dolphin Cotylocara is a xenorophid and exhibits
a series of sinuses relating to echolocation, showing that xenorophids could
produce high frequency sounds. This periotic reported by Travis Park et al.
from the upper Oligocene Belgrade Formation of North Carolina has a loosely
coiled cochlea and other features indicative of high frequency hearing -
leading these authors to propose that echolocation was an adaptation present in
the earliest odontocetes. See Churchill et al., above, which builds
significantly on this study.

Peredo, C.M. and M.D. Uhen. 206. A new basal chaeomysticete
(Mammalia: Cetacea) from the late Oligocene Pysht Formation of Washington, USA.
Papers in Palaeontology 2:4:533-554.
http://onlinelibrary.wiley.com/doi/10.1002/spp2.1051/abstract
As outlined above, eomysticetids are the earliest diverging
functionally toothless baleen whales. Eomysticetids are typically mid-late
Oligocene in age, have elongate mostly toothless rostra, large jaw muscle
attachments, earbones intermediate between archaeocetes and Miocene mysticetes,
and archaic basilosaurid-like postcranial skeletons. They've been reported from
South Carolina, Austria, Japan, and New Zealand; an occurrence in Japan
suggests we should be finding them in the eastern North Pacific. This study
reports an eomysticetid-like baleen whale, Sitsqwayk cornishorum,
from the upper Oligocene Pysht Formation of the Olympic Peninsula (formerly
known for the small aetiocetid Fucaia goedertorum). Sitsqwayk has a few
key differences from published eomysticetids, including a wider intertemporal
region and the lack of a sagittal crest. However, Sitsqwayk does possess
at least two eomysticetid synapomorphies – a zygomatic process that lacks a
supramastoid crest and is longitudinally rotated so that the lateral surface
faces dorsolaterally. Sitsqwayk showed up on a cladogram one node below
Eomysticetidae. I imagine that with discovery of better earbones and a skull
with complete nasals, eomysticetid relationships may become more firm.
Regardless, Sitsqwayk represents the first named Oligocene
chaeomysticete from the eastern North Pacific and I heartily welcome additional
finds to be published! Similar undescribed skulls exist in collections from the
Oligocene of New Zealand, and perhaps species like Whakakai waipata may
be somewhat related.

Until recently the fossil
record of marine mammals was generally quite localized and consisted of
published occurrences that were either 1) outdated and improperly identified or
2) spotty and too incomplete to reveal any patterns of biogeography. Recent
advances have published assemblages in their entirety – not just focusing on
the best preserved material, and instead giving a more honest view of the fauna
– refined old identifications – and reported new specimens and taxonomic
occurrences. In the last decade, a number of studies have taken advantage ot
the rapidly improving published record and begun broad-scale analyses of marine
mammal distribution, biogeography, dispersal, evolution, as well as basic
sampling problems. A classic biogeographic hypothesis in marine mammal
paleontology is the use of the central American seaway – open until 3 million
years ago prior to closure by uplift of Panama
– whether walruses, baleen whales, seals, sea cows, or various dolphins. This
new study uses the paleobiology database to statistically analyze patterns of
faunal similarity between different ocean basins. Notable findings include 1)
broad Eocene similarity between the North Atlantic and Mediterranean/Tethys, 2)
faunal differentiation between basins driven by the Messinian Salinity Crisis
at the end of the Miocene, and 3) faunal similarity between the Pacific and
Atlantic pretty much only during the middle Miocene. Perhaps most
significantly, this study suggests that the Central American Seaway was a much
less used corridor for dispersal – and many marine mammals which inhabit the
North Pacific and North Atlantic instead likely dispersed through the Arctic
instead, which opened up around the time the CAS closed. This is not the first
time this has been proposed, as it was originally proposed by Kohno et al.
(1995) for tusked walruses and later by Lambert (2008) for phocoenid porpoises;
I discussed this idea at some length in my 2013 Geodiversitas monograph, in
addition to Pliocene faunal provinciality (e.g. dissimilarity). The current
study is the first to provide a semi-quantitative statistical demonstration of
these ideas, which feels quite vindicating!

Baleen whales like the blue whale (Balaenoptera musculus)
are the largest vertebrates of all time (sorry sauropod researchers). Living in
water over terrestrial settings has its benefits – energetically more efficient
locomotion thanks to buoyancy, and generally more abundant food. The lower
constraints on on biomechanics have permitted gigantism – which in turn allows
for greater capacity for long distance migration and dispersal; it's no
coincidence that marine mammals with the largest body sizes have some of the
largest ranges as well (in stark contrast to tiny seabirds like terns, which
have similar migration distances yet weigh only a few ounces). In marine
mammals, body size likely reflects niche as well as food availability- the
latter of which is controlled in a “bottom up” fashion by primary productivity
(the amount of chemical energy as biomass that primary producers – e.g.
plankton – produce through photosynthesis or similar means). This study tracks
maximum body size in in two different groups: filter feeding marine mammals
(Mysticeti) and herbivorous marine mammals (Sirenia, Desmostylia) throughout
the Eocene-Recent. Curiously, baleen whales do show much smaller sizes
throughout most of their fossil record, attaining a large size only in the
Plio-Pleistocene interval; similarly, minimum body size for baleen whales was
substantially lower during the Oligocene and Miocene. This is demonstrated for
the second time – a pattern already discovered and acknowledged by Olivier
Lambert and colleagues (2010: Nature, Livyatan melvillei paper). This
suggests that mysticete gigantism corresponds to Pleistocene-age increase in
primary productivity driven by intensified chemical weathering. On the other
hand, most sirenians and desmostylians show a comparably stable trend in body
size, with the hydrodamalines (the western sea cow Dusisiren, and the
extinct giant sea cows Hydrodamalis) – coinciding with the late
Miocene-Recent diversification of kelps and proliferation of kelp forests. For
more analytical examinations of body size trends in other marine mammals, be
sure to check out Morgan Churchill et al.'s paper on pinniped body size and
Cope's Rule.

These short papers are a comment and reply on Crerar et al.
(2014), a paper proposing that a second
population of Steller's sea cow (Hydrodamalis gigas) inhabited the
shoreline of Saint Lawrence Island, located in the northern Bering Sea over a
thousand miles to the northeast of where the first sea cows were discovered
(Commander/Komandorskiye Islands) and subsequently hunted to extinction in the
mid 18th century. The original paper apparently used a number of
specimens of “mermaid ivory” - the bones of the Steller's sea cow are dense
enough for carving and polishing by native artists like regular ivory (thick
teeth/tusks). These suspected Hydrodamalis bone carvings were acquired
by the authors from ivory carvers and artists in Alaska, sampled for molecular
analyses, and radiocarbon dated. However, the collection to which the physical
bone samples are simply listed as “Crerar” - in other words, the private collection
of the lead author. Pyenson et al. point out that the specimens should have
been preserved as voucher specimens in an established museum collection – and
since this was not done, the morphological identification of the specimens is
essentially untestable. They further point out that serious ethical concerns
are associated with the purchase and study of specimens from fossil dealers –
outlined in more detail by the Society of Vertebrate Paleontology Ethics
Statement. Crerar et al. respond by highlighting the significance of their
discoveries, as well as indicating that the samples had been acquired as part
of an investigation into illegal trade of other marine mammal remains – but
because Hydrodamalis is extinct, it is exempt from restrictions imposed
by CITES, US Endangered Species Act, or the Marine Mammal Protection Act. I
agree that all publications involving fossils absolutely must rely on specimens in museum
collections, but there are plenty of papers out there reporting purchased
specimens that ended up in museum collections (e.g. Marx and Kohno, above). There is a bit of grey area which in all likelihood is never going to go away.

It's been a good year for
the study of cetacean earbones and hearing using CT data – see for example
Ekdale, Churchill et al., and Park et al. (above). These analyses have
effectively made the destructive and time-consuming practice of serial grinding
completely obsolete. The lead author, Dr. Racicot, CT scanned the pterygoid
sinuses of porpoises as part of her master's thesis at San
Diego State (now
published), plunging her into the study of porpoise anatomy and evolution and
including the description of the bizarre porpoise Semirostrum ceruttii.
This study originated during her Ph.D. and benefited from data collection in Japan.
This study presents new scan data and an analysis of the bony labyrinths of
extinct and extant true porpoises (Phocoenidae). Unsurprisingly, all extinct
phocoenids exhibit features consistent with high-frequency hearing. This
analysis identifies characteristics permitting inference of an extinct
odontocete's sensitivity to head movements – with modern pelagic phocoenids
having a high sensitivity. The extinct, presumed benthic feeding porpoise
Semirostrum had low sensitivity to head movements – consistent with life on the
continental shelf. This study furthermore proposes a method to infer body size
from measurements of the labyrinth. However, body length estimates of 7 meters
for two Japanese fossil phocoenids – Haborophocoena and Numataphocoena
– seems unbelieveably long as the skulls are not quite that large. Perhaps this
is a result of highly derived bony labyrinths and some unknown specialization
in these species. Regardless, the promise for estimating body size from an
isolated earbone is tantalizing.

The true seals (Phocidae) are one of the more challenging
groups of pinnipeds to study in the fossil record. The majority of the fossil
record of phocid seals consists of isolated postcrania - aside from scattered
mandibles and fragmentary skulls and a few skeletons from South
America (Acrophoca, Hadrokirus, Kawas, Piscophoca). The
fossil record of phocids from Europe and western Asia is particularly
difficult, being almost completely based on postcrania with confusing taxonomic
history. One of the few early phocid taxa represented by well-preserved crania
is Devinophoca - two species within this genus have been reported from
the same quarry in Slovakia. Both species (Devinophoca claytoni, Devinophoca
emryi) are known from nearly complete skulls differing in only a few minor
details - some of which appear to be incorrectly diagnosed (for example, the
infraorbital foramen of D. claytoni is claimed to be smaller than D.
emryi, yet the converse appears to be true). Devinophoca emryi
was named in 2015, and I'm skeptical that it represents a second species.
Regardless of how many species are recognized, the Devinophoca material
from the "Bonanza" quarry clearly represents an early phocid. This
study refers various postcranial elements to Devinophoca emryi,
including scapula, humerus, radius, ulna, innominate, femur, tibia, fibula,
calcaneum, and astragalus. Some of the postcranial features are clearly
phocid-like (e.g. laterally everted innominate, posterior extension of
astragalus limiting motion of ankle) yet other bones are remarkably similar to
the early pinnipedimorphs Enaliarctos and Pinnarctidion (e.g.
humerus). The isolated bones are referred to D. emryi over D.
claytoni owing to the "Ecomorphotype" method of Koretsky (2001);
modern species occupying different niches have specific morphologies of the
cranium, mandible, and postcrania corresponding to their niche. This is taken a
step further, and isolated postcrania are referred to taxa with
craniomandibular material based on shared "ecomorphology" by analogy
with extant taxa. Unfortunately, this method is perhaps circular reasoning at
best and untestable at worst. For example - these authors explain that, based
on the different ecomorphotype groups outlined by Koretsky (2001), the mandible
and postcrania of D. emryi clearly represent a single taxon - but they
never explain why the postcrania do not belong to D. claytoni, which is
not known by a mandible. Because the ecomorphotype method focuses on mandibles
and postcrania and not skulls, the question of whether or not this material
represents D. claytoni is simply left unaddressed.

As mentioned several times
earlier, this has been a bit of a ridiculous year for advances in beaked whale
paleontology – with the description of Chavinziphius and Chimuziphius
(Bianucci et al.), new specimens of Messapicetus (Bianucci et al.),
histology of the bizarre rostrum of Globicetus (Dumont et al.), and a
new species of Africanacetus dredged from the Atlantic off Brazil
(Ichishima et al.). This new study reports a new genus and species of beaked
whale, Dagonodum mojnum, from the upper Miocene Gram Formation of
Denmark. The Gram Formation has previously yielded the holotype skulls of the
“kelloggithere” mysticetes Uranocetus gramensis and Tranatocetus
argillarius, and some poorly preserved pontoporiid dolphin skulls. Dagonodum
is a reference to the Lovecraftian sea god Dagon (which is hilarious, given a
conversation with colleagues Brian Beatty and Morgan Churchill about how
awesome it would be to name an extinct whale after Dagon). Dagonocetus
is similar to Messapicetus and has a pair of enlarged mandibular tusks
and a polydont, homodont dentition set in an elongate narrow rostrum. Enlarged
tusks further identify the holotype as a male. The type also includes a decent
pair of earbones. Dagonodum was likely not as specialized for suction
feeding as modern mostly-toothless ziphiids, and furthermore possessed somewhat
more elongate cervical vertebrae indicating a higher degree of neck flexibility
than modern ziphiids.

Image by Gabe Santos
Desmostylians are a bizarre group of hippo-like marine
mammals that are closely related to elephants (Proboscidea) and sea cows
(Sirenia) and are within the Afrotheria (African mammals). The earliest
desmostylians are Oligocene in age (see Beatty and Cockburn, above) and fossils
of Desmostylus are some of the more common occurrences in the Miocene of
California, Oregon, and Japan. Desmostylians have highly derived teeth – tusk
like canines and incisors, and premolars and molars with cusps arranged into
finger-like tubes of enamel that superficially resemble six-packs of beer cans.
Much attention has been afforded to the taxonomy and functional anatomy of
desmostylians, but little research on the ontogeny and tooth replacement of
these strange mammals has been undertaken. This study reports an enormous
mandible of Desmostylus hesperus from the mid-upper Miocene Sespe
Formation of Orange County, CA. This mandible is quite a bit larger than all
previously published specimens of Desmostylus, and uniquely has a
downturned lower tusk, resembling the strange giant elephant Deinotherium.
Of greater importance is the lack of teeth – though a large bony dental capsule
is present as a large knob on the inside of the mandible. This suggests that Desmostylus
continued to thrive after loss of its last pair of molars – and perhaps fed
on kelp in old age. Kelp may be consumed without teeth, as the extinct
Steller's sea cow Hydrodamalis gigas fed solely on kelp without having
any teeth. The extreme size of this specimen, along with the absence of cheek
teeth and infilling of alveoli – indicates that Desmostylus had a dental
eruption pattern most similar to other afrotheres, where the last molar is not
erupted until the mandible attains full size. A recent study suggests that
desmostylians may be early diverging perissodactyls (odd toed ungulates);
either the the dental eruption pattern means that it is due to shared ancestry,
or that this dental eruption pattern evolved twice – once within Afrotheria and
once within Perissodactyla. As a side note, I saw this spectacular specimen in
2013 when I visited the Cooper Center and because of its gigantic size, I
thought it was some weird elephant like a gomphothere – to which Gabe Santos
was delighted to inform me that it was a giant Desmostylus the size of a
small elephant.
These two new studies report newly discovered specimens of
the porpoise Numataphocoena from the Pliocene of Japan. Porpoises are
currently distributed worldwide but now known to have been restricted to the
Pacific for most of their evolution, with diverse Mio-Pliocene assemblages now
known from Japan,
California, and south America. Numataphocoena
was originally described from a partial skeleton (including a fragmentary
skull with well-preserved earbones and a nice postcranial skeleton) from the
early Pliocene of Hokkaido. Despite cranial differences, Numataphocoena
already had the hallmark "spatulate" teeth of modern porpoises - most
modern odontocetes have conical teeth. A new periotic reported in the first of
the new papers suggests closer affinities with another Pliocene porpoise from
Hokkaido, Haborophocoena. The referred periotic further illuminates some
subtle ontogenetic trends in odontocete earbones - an understudied topic in my
opinion. The second new paper (Tanaka and Ichishima) report a new cranium of Numataphocoena,
including a nearly complete braincase but incomplete rostrum. Similar to the
prior paper, this specimen is ontogenetically younger and reveals some
information about ontogeny in extinct porpoises, and more critically expands
the known morphology of the species. Cladistic analysis supports a novel clade
of phocoenids including practically every porpoise from Hokkaido from the
latest Miocene and early Pliocene: Haborophocoena spp., Numataphocoena,
Archaeophocaena, and Miophocaena. I've long suspected that
Japanese phocoenids are oversplit, and this grouping may suggest that the
number of species and genera ought to be trimmed.

The earliest Miocene (Aquitanian) is a period in time with
abundant marine rocks but rare fossil marine mammals. Some of the only areas
with earliest Miocene marine mammals include California
and Oregon (e.g. Jewett Sand,
Skooner Gulch Formation, Nye Mudstone), Italy
(Belluno Sandstone), and New Zealand
(Milburn Limestone, Mt. Harris Formation). Quite a few fossil odontocetes
including complete skulls from the Belluno Sandstone of Italy have been
reported – however, dating of this unit is poor and the unit possibly
Burdigalian. The Belluno Sandstone assemblage is dominated by longirostrine
odontocetes of various families (Dalpiazinidae, Eoplatanistidae,
Eurhinodelphinidae, Squalodelphinidae), sperm whales, and squalodontids – and
notably lacks baleen whales. New Zealand has offered a few earliest Miocene
cetaceans with good dates – including the early “kelloggithere” Mauicetus
parki, the squalodontid “Prosqualodon” hamiltoni, and the recently
named waipatiid-like dolphin Papahu taitapu. The holotype of Papahu was
collected from Nelson (northwestern part of the South Island, NZ), and includes
a nearly complete skull with partial periotic, and several vertebrae including
the axis. Originally suspected to be a kentriodontid dolphin, cladistic
analysis suggested Papahu was further down on the odontocete tree, and
well outside crown Odontoceti. This new paper reports new material of Papahu
– a specimen identified as cf. Papahu sp., including a fragmentary
skull with a more completely preserved periotic and a complete tympanic bulla.
Inclusion of the new specimen pulls Papahu into two different
phylogenetic positions: either as an early diverging member of Platanistoidea
(see Boersma and Pyenson, above) or as the earliest member of Synrhina
(Delphinida + Ziphiidae). Regardless – this new specimen and analysis strongly
suggests that Papahu is indeed a crown odontocete, and highlights the
utility of fragmentary specimens.

This paper marks one of the last entries in Yoshi Tanaka's
Ph.D. thesis on Oligocene odontocetes from New Zealand. Earlier papers from his
thesis reevaluated "Prosqualodon" marplesi and reassigned
it to the new genus Otekaikea, named the new species Otekaikea huata,
and redescribed "Microcetus" hectori and transferred it
to Waipatia. Waipatiid dolphins are generalized heterodont odontocetes
from the Oligocene and likely early Miocene, likely filling in a bottlenose
dolphin-like niche - though waipatiids notably differ from later groups of
odontocetes in having moderately enlarged tusk-like incisors. This new study
reports the new genus and species Awamokoa tokarahi, a new waipatiid
sister to Waipatia spp. This new species differs from Waipatia spp.
in a few features of the basicranium and periotic. Awamokoa is
represented by a partial skull, well-preserved earbones, a partial mandible,
various teeth, vertebrae, ribs, partial scapulae, ulna, and phalanges. Awamokoa
represents the fifth confirmed Oligocene waipatiid from New Zealand (Papahu
may represent a sixth) and one of the best examples of waipatiids preserved
with postcrania; other waipatiids are likely to be reported from elsewhere
(e.g. see Fitzgerald, above). Awamokoa has a relatively shallow and
elongate temporal fossa, suggesting that the feeding apparatus was adapted for
slower but more powerful bites, and that later diverging platanistoids
(squalodelphinids, Platanista) with anteroposteriorly shorter and deeper
temporal fossa housed muscles adapted for rapid snapping of the lower jaws.
This transition similarly parallels the development of homodonty within the
Platanistoidea. Lastly, Awamokoa is from the early late Oligocene
Kokoamu Greensand, and approximately 28-25.2 Ma in age - making Awamokoa
one of the earliest occurring platanistoids. Arktocara yakataga,
an allodelphinid (see Boersma and Pyenson, above) may be older but has poorer
age resolution. Furthermore, waipatiids may not be within the odontocete crown
and instead may not be related to true platanistoids (e.g. Squalodelphinidae +
Platanistoidea) - see various phylogenies published by Jonathan Geisler and
colleagues.

Not all whale specimens are spectacular, with completely
preserved skulls, mandibles, and postcranial skeletons. In fact, owing to the
large size of baleen whales in particular, most are rather incomplete; it's
difficult to catch a large skull eroding out and nab the whole thing. Earbones
are often fairly informative, and in the case of a new species reported in this
study, Whakakai waipata, well-preserved earbones clearly demonstrate
that this baleen whale is a new genus and species; the earbones are distinctive
and well-preserved enough that they serve as the major features that diagnose
this new species. Whakakai is from the upper Oligocene Kokoamu Greensand
of New Zealand, collected nearby the type localities of the odontocete Waipatia
maerewhenua and the eomysticetid Tohoraata raekohao, though from
somewhat stratigraphically lower than each. The preserved portion of the skull
indicates that this mysticete lacked a sagittal crest – immediately
distinguishing it from eomysticetids, but resembling such mysticetes further up
the tree as Horopeta and Mauicetus, both from the overlying
Otekaike Limestone. The earbones on the other hand are quite strange. The
periotic is enormous and inflated, with proportions somewhat resembling other
archaic mysticetes – yet has a relatively small pars cochlearis; the posterior
process is unfused, like other archaic mysticetes. The tympanic bulla is
eomysticetid-like with well-defined medial and lateral lobes, but has a
partially closed elliptical foramen – which is completely closed in later diverging
mysticetes (Balaenomorpha). Cladistic analysis demonstrates that Whakakai
is more derived than eomysticetids and plots further up the tree – close to Horopeta
and Mauicetus, and in one of two analyses, as sister to Horopeta (also
named in a paper resulting from C-H Tsai's thesis at U. Otago. These species
evidently lived alongside the more primitive eomysticetids, yet illustrate a
suite of morphologies broadly intermediate between eomysticetids and Miocene
“kelloggitheres”.

Perhaps one of the strangest and more obscure extinct marine
mammals is Kolponomos - an ursid-like semiaquatic carnivore with
enlarged, sea otter-like teeth, binocular vision, and enlarged procumbent
canines. Originally discovered from the lowermost Miocene Clallam Formation of
Washington in the 1960s and thought to be a giant marine raccoon, additional
well-preserved fossils including crania were reported in the 1990s and revealed
ursid affinities, and a possible sister taxon relationship with pinnipeds (I
suspect that Kolponomos along with Potamotherium and Puijila represents
an aquatic diversification of arctoid carnivores related to pinnipeds). The
dentition of Kolponomos was inferred to be adapted for crushing
mollusks, and enlarged mastoid processes were interpreted as anchoring strong
neck muscles. This new study investigates the feeding morphology of Kolponomos
using finite element modeling (FEM). This study notably looks beyond arctoid
carnivores for analogies, and found that the deep "chin" of the
mandible shares certain biomechanical similarities with the sabertooth cat Smilodon.
In carnivorous mammals that chew on hard food, there is a tradeoff between
"stiffness" and mechanical efficiency: a stiff mandible does not
store much energy, and a more flexible mandible stores energy during a bite.
Based on analogy with Smilodon, Kolponomos was hypothesized to
possess high mandibular stiffness - which was borne out by the analysis.
Surprisingly, though similar in feeding morphology, the sea otter possessed the
most flexible mandible. Kolponomos is inferred to have utilized an
"anchor bite" stage prior to chewing, where the mandible and lower
dentition helped pry mollusks from rocks, followed by otter-like crushing of
the mollusk with the large cheek teeth. Enlarged neck muscles gave Kolponomos
the ability to apply enormous torque onto attached mollusks (e.g. mussels).
This study indicates that Kolponomos and the sea otter Enhydra
crush hard shelled prey while independently relying on mandibular stiffness and
high bite force (respectively), using somewhat different pathways which have
nonetheless resulted in craniodental convergence. Because higher stiffness is
required for the anchor bite, I imagine that the lower stiffness in Enhydra is
permitted by the fact that Enhydra removes mollusks and crustaceans from
the substrate by using its forelimbs rather than its feeding apparatus.

If you've read everything above you're already familiar with
the utility of isolated cetacean earbones – they can help us diagnose species,
inform us about the hearing ability of extinct cetaceans, intraspecific
variation, and the diversity of extinct cetacean faunas. The Yorktown Formation
at the Lee Creek Mine in North Carolina has yielded one of the largest Pliocene
marine mammal assemblages in the world, yet the cetacean assemblage is
dominated by isolated (yet diagnostic) earbones. Particularly common earbones
include delphinid dolphins and small physeteroid sperm whales. In 2008, the Lee
Creek IV volume was finally published after being in press and in preparation
for decades, and reported the presence of large periotics of some sort of
unknown kogiid (pygmy sperm whale). These earbones are much larger than extant Kogia
– and this study reports the first records of periotics of true Kogia,
as well as additional specimens of the larger species. These same large kogiid
periotics are also reported from the Bone Valley Formation of Florida, the
marine mammal assemblage of which is mostly undescribed. A large kogiid, Aprixokogia
kelloggi, was named in the Lee Creek IV volume yet the skull did not
include earbones. It is possible that these large periotics are attributable to
Aprixokogia, but skulls with associated periotics are needed to be sure.
What is troubling is that a recent donation to our museum (CCNHM) from the mine
includes two fragmentary kogiid skulls – one of which appears to be Aprixokogia,
and the other is a second non-Kogia kogiid whale – indicating the likely
presence of three kogiids, though only two appear to be represented by
periotics.