
Within the world of paleontology there
is on occasion a disconnect between the names, ideas, and concepts
embraced by academics and amateur fossil collectors. Some of us
academics do our best to educate the community, and others make
little attempt. The purpose of this post is to clarify several
commonly believed ideas I've heard amongst fossil collectors, but
evidently have either been discounted within the peer reviewed
literature or originated completely outside the world of academic
paleontology. I jokingly refer to these sorts of ideas as "fossil
folklore"; a very, very common version of this is "I knew a
guy who found an 8 inch long megalodon tooth" (they
rarely exceed 6.5 inches and do not really exceed 7) and every
Miocene shark tooth locality I've ever been to has an apocryphal
story told by a guy who's visited the site for decades, only to have
a perfect 6" C. megalodon tooth be found by a kid after
only a few minutes of searching. This post deals with various marine
mammal related "folklore" I've heard, and hope to set the
record straight.
No, you don't have a Prosqualodon
tooth
This first myth applies principally to
North America, and didn't really bother me until I had spent a few
years in the field of marine mammal paleontology. Fossil collectors
in the US often refer to large odontocete (toothed whale) teeth that
do not match Squalodon (see below) as Prosqualodon.
Most of these teeth look nothing like Squalodon - they are
single rooted, with a greatly inflated root, and a small conical
crown. I guarantee that if you search "Prosqualodon tooth"
on google you will see dozens of these for sale. One site goes so far
as to call these "Prosqualodon errabundus", which is
extra hilarious - and I'll explain why shortly. It seems as though
the thought process follows a simple pattern like this: if an
odontocete tooth is small, then it is a "porpoise" (see
below as well) or "dolphin" and no further attempt at
identification is made. If it is a very large tooth with an inflated
root and a small crown (or completely worn away crown), then it is a
sperm whale. If the tooth is large but double rooted, triangular, and
cuspate, then it is Squalodon. And lastly, if the tooth is
Squalodon-sized but single rooted with a small conical crown
and an inflated root but too small to be a "sperm whale",
then it is Prosqualodon. This identification process is
obviously oversimplified, but there's a bigger and even more
glaringly obvious problem here: Prosqualodon has never been
reported in North America.

A large odontocete tooth with a thickened neck of cementum from the Sharktooth Hill Bonebed (photo from www.thefossilforum.com) that are typically identified as "Prosqualodon" or "Prosqualodon errabundus" by collectors and fossil dealers.
Several species of Prosqualodon
have been reported, but all from the southern hemisphere. The
type species is Prosqualodon australis, described in the late
19th century and now known from several skulls from the lower Miocene
of Patagonia - and morphology is consistent with this taxon being a
short-snouted squalodontid. Later in the 20th century, three species
were named from Australia and New Zealand. Prosqualodon
davidis was described from the lower Miocene of Tasmania, and
is arguably the best known of all the four species - with a nearly
complete dentition, complete skull, partial mandibles, vertebrae, and
an articulated flipper. Unfortunately, the skull is lost but it was
figured beautifully by Flynn (1948). Prosqualodon
davidis is very similar (also short-snouted), and clearly
closely related to P. australis. Prosqualodon marplesi
has had a rocky taxonomic history - originally reported by Professor
Benham (former zoology chair at my doctoral alma mater, U. Otago),
who was a bit arrogant yet committed various taxonomic gaffes during
his career. Prosqualodon marplesi was based on an incomplete
skull, but transferred to the genus Notocetus within the
Squalodelphinidae by Fordyce (1994) based on dental and braincase
features as well as cladistic results, and then reassigned to the new
genus Otekaikea and placed within the Waipatiidae by Tanaka
and Fordyce (2014). The very last species, Prosqualodon
hamiltoni, is likely a squalodontid (Fordyce, 1994) but
probably a long-snouted form and the skull is not particularly
similar to Prosqualodon australis or P. davidis
(Fordyce, 1991). Regardless of these New Zealand species being
removed from Prosqualodon, something close to Prosqualodon
davidis is recorded from the lower Miocene Mt. Harris Formation
of New Zealand (Fordyce and Roberts, 2009) but has yet to be
described. The only other reported occurrence of Prosqualodon is
a couple of vertebrae from the lower Miocene of Venezuela
(Sanchez-Villagra et al., 2000) - perhaps not such a reliable
occurrence, but technically the only one from the Northern
Hemisphere.


The skull and teeth of Prosqualodon davidis from the earliest Miocene of Tasmania. Note the lack of cementum and the complex crown with rugose enamel and accessory cusps - nothing like the North American specimens. From Flynn (1948).
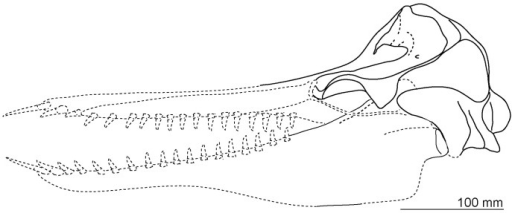
The skull of the dolphin formerly known as Prosqualodon - now Otekaikea.
From Tanaka and Fordyce (2014).
No fossils of true Prosqualodon
have been reported from North America, nor have I seen any
unpublished specimens in museum or private collections. Further to
the point - why aren't these fossils widely called "Prosqualodon"
by US collectors actually Prosqualodon? Prosqualodon
davidis has teeth that are similar in most respects to other
squalodontids - conical anterior teeth with fluted enamel and double
rooted cheek teeth with accessory cusps and finely rugose enamel.
Teeth of "Prosqualodon" from the US have nothing in
common with true Prosqualodon other than having enamel and a
root. So, if not Prosqualodon, then what are they? Could they
be "Prosqualodon errabundus" as some commercial
fossil dealers call them?
"Squalodon"
errabundus was originally reported by Kellogg (1931) off of a
single periotic (inner ear bone) from the Sharktooth Hill Bonebed of
California. Kellogg considered the periotic to be broken and similar
to Squalodon, but it would not be until the last few years that the
true affinities of "Squalodon" errabundus
would be uncovered. More complete material indicates that "Squalodon"
errabundus is actually a strange long snouted "platanistoid"
dolphin in the extinct family Allodelphinidae, currently known only
from the early and middle Miocene of California, Oregon, Washington,
and Japan. I've seen references to "Prosqualodon
errabundus" for over a decade, though this binomial
combination has never ever been proposed once in the literature - and
the idea that this species is not a squalodontid has been kicking
around since the mid 1970s, though only formally transferred to
Allodelphinidae recently. In 2009, "Squalodon"
errabundus was given a new genus and is now Zarhinocetus -
seven years ago now, and yet you can still find teeth for sale as
"Prosqualodon errabundus". The fact that the
species has had its own genus now for almost a decade makes the
"Prosqualodon" issue even more humorous.
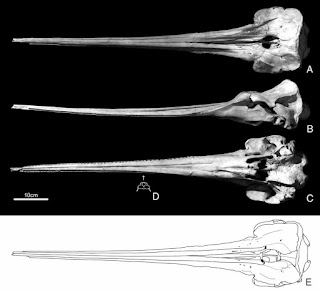

The skull of Zarhinocetus errabundus from the Sharktooth Hill Bonebed - note the tiny tooth sockets. From Kimura and Barnes (2016).
Which brings us full-circle back to
"Prosqualodon" fossils from the US. Could they be
Zarhinocetus errabundus? First of all, many "Prosqualodon"
teeth are reported from the east coast, and allodelphinids are
unknown outside the Pacific. We don't actually know what the teeth of
Zarhinocetus errabundus look like, because a tooth has never
been found in a jaw or skull condifently identified as Z.
errabundus. However, amusingly enough, the tooth sockets of
Zarhinocetus are tiny and much too small to house the large
teeth commonly identified as "Prosqualodon", and the
teeth of closely related Allodelphis and Ninjadelphis are
tiny needle-like teeth only 1.5 mm wide at the base. Truth be told,
the distinctive specimens from Sharktooth Hill belong to a completely
unidentified odontocete with extensive cementum deposits on the
roots, leading to enormous inflated roots that dwarf the crown. Aside
from this, these teeth are reminiscent of delphinoid teeth - and in
my opinion, are similar to teeth of the giant kentriodontid dolphin
Hadrodelphis calvertensis from the Calvert Formation of
Maryland. They are all single rooted, but decidedly more massive;
based on giant earbones I've seen, a giant kentriodontid dolphin is
lurking in the Sharktooth Hill Bonebed, waiting to be described - and
may be the "owner" of these teeth. In summary,
identifications of fossil teeth should be based on anatomical
comparisons and not hearsay - which is arguably a failure of
professional paleontologists not interfacing enough with the amateur
community to set some of these things straight.


A maxilla fragment with teeth (figured upside down as it was originally misidentified as a mandible) of Squalodon sp. from the Miocene of New Jersey. Specimens of true Squalodon from the Atlantic Coastal Plain are restricted to Miocene rocks. Image from https://vmnhpaleontology.wordpress.com.
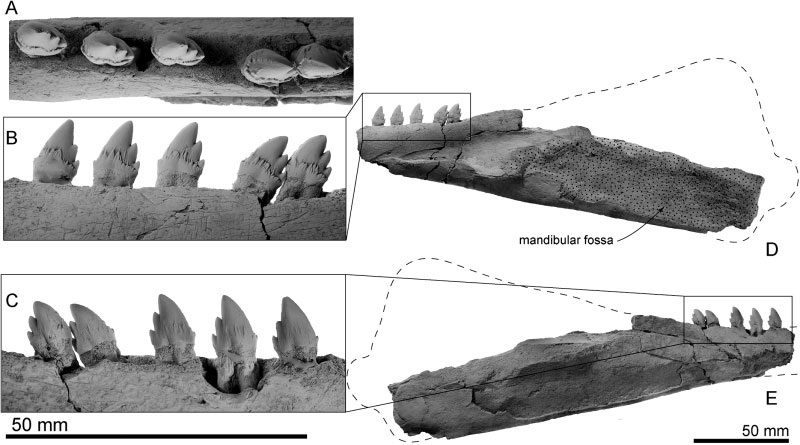
Not Squalodon: Waipatia hectori, Waipatiidae, Oligocene, New Zealand; very similar teeth are found in the Charleston area. From Tanaka and Fordyce (2015).

Also not Squalodon: Echovenator sandersi, Oligocene, Charleston, South Carolina. From Churchill et al., (2016).
Do you really have a Squalodon
tooth?
Odds are, unless you're collecting in
the Calvert Formation in Maryland/Virginia or the Pungo River
Limestone in North Carolina (as far as North America is concerned
anyway), you have probably not found a Squalodon tooth. Why do
I say this? All bona fide records of Squalodon in the USA come
from those two rock units. There are a couple of random records from
New Jersey (e.g. the Squalodon atlanticus holotype).
South Carolina, despite popular belief to the contrary, does not have
any actual specimens of Squalodon that are verifiable. There
is Eosqualodon (formerly identified only as Genus X) from the
Oligocene Chandler Bridge Formation, which is similar in size to the
smaller Calvert species Squalodon calvertensis, but
has teeth that are somewhat more denticulate.
So why identify something as
Squalodon? What makes Squalodon? In my own experience
collectors assume that any cetacean tooth that resembles a tiny
archaeocete tooth with double-roots and accessory cusps is Squalodon.
True, Squalodon is one of the last truly heterodont
odontocetes - heterodonty is the primitive condition where cheek
teeth and incisors look very different. But is Squalodon the
only heterodont odontocete? As it turns out, nearly every group of
dolphins in the Oligocene were heterodont - including agorophiids
(common in the Oligocene of South Carolina), simocetids (possibly
present), waipatiids (also common), and xenorophids (perhaps the most
common odontocetes). There's even a Squalodon-like giant
dolphin known currently only as "Genus Y" that has
heterodont teeth, but may not be closely related to squalodontids at
all.
Collectors in the Charleston area
should, for the time being, stop identifying local specimens as
Squalodon, since I've not seen any from this area that can be
identified as Squalodon - we only have one real South Carolina
Squalodon specimen in our collections at CCNHM, but it's a
tooth found down in Beaufort County where there are Miocene rocks
exposed.
Carcharodon hastalis teeth found in association with a skeleton of the Miocene baleen whale Incakujira from Peru (Yuji, 2014).
“I found this shark tooth near
this whale bone – the shark must have eaten it!”
This is a very common misconception.
Whale bones are often found with shark tooth bite marks – a
skeleton recently donated to us has bite marks on more than 30% of
the skelton, and relative to the paltry number of such traces I
recorded in the Purisima Formation assemblage, the number of
shark-bitten bones on the east coast is staggering. Such traces are
god evidence of fossil behavior – and different traces reflecting
different types of teeth have been named as trace fossils
(ichnofossils). However, the case is more difficult to make when
there are no such traces. Slow deposition can result in a slightly
higher than normal concentration of teeth in addition to marine
mammal bones. Association of bones and teeth – especially if the
“skeleton” consists of just a couple of bones or a single bone –
are commonly formed by sedimentary reworking and concentration. If
any sign of transport or reworking is obvious – abrasion, polish,
fragmentation – then the fossil association is almost certainly
caused by reworking. If the fossils were found in a bonebed, then
it's not possible to infer any behavior. The following conditions
must be met to infer predation-based association of shark teeth and a
prey item in the marine vertebrate fossil record: 1) there must be
evidence that the prey animal (marine mammal in our case) represents
a single individual and is isolated in the sediment; 2) there must
not be any evidence that the associated teeth are from a condensed
section, during which vertical differences may not indicate closeness
in time; 3) the shark teeth must be found near the skeleton and in a
higher concentration than surrounding sediment. Classic bulletproof
examples include articulated marine mammals and marine reptiles
preserved in fine-grained deposits with dozens to hundreds of shark
teeth clustered in a halo around the skeleton. In extremely rare cases shark teeth may even be embedded into bone - clear evidence of predation activity.


Some baleen whale vertebrae with unfused vertebral epiphyses - Miocene Calvert Formation, Carmel Church Quarry, Virgina. Image from https://vmnhpaleontology.files.wordpress.com.
“There are a lot of unfused
vertebrae here – so this was a whale calving ground.”
Isolated cetacean vertebrae are
informally considered some of the most boring fossils from Miocene
and Pliocene rocks. Whales and dolphins have high vertebral count,
and owing to the loss of hindlimbs, whale and dolphin skeletons are ¾
ribs and vertebrae by count. Fusion of epiphyseal plates – whether
in vertebrae, or limb bones – is a hallmark feature of mammalian
growth. These are the so-called “growth plates” that doctors will
evaluate growth/cessation of growth in young teens based on X-rays.
Several basins, including the Chesapeake embayment (e.g. Calvert
Cliffs) and the Charleston embayment (Oligocene Ashley/Chandler
Bridge formations) have been formally and informally identified as
“calving grounds” owing to the high number of cetacean vertebrae
with unfused epiphyses. However, studies of modern cetacean
epiphyseal demonstrate that not all vertebrae in the column fuse in
every species, and it does not necessarily correspond to maturity.
Furthermore, epiphyseal fusion occurs when cetaceans (like other
mammals) occurs relatively close to skeletal maturity – so
cetaceans will have unfused vertebrae for a period of several years.
This means that individuals significantly older than neonates
(calves) will possess unfused vertebrae, and thus the presence of
abundant unfused vertebrae in a particular area has no bearing on age
as it could signify somewhat older individuals that have already
undertaken migrations (and may have been calved/born in a completely
different region). Some species retain unfused vertebrae longer than
others, so there is a taxonomic complication as well.

Conical teeth in a dolphin (Delphinidae, left) and spatulate teeth in a porpoise (Phocoenidae, right). Image from www.http://wildwhales.org
"This tooth is from a dolphin
and this one is from a porpoise."
This is a common one. Truth be told,
when fossil collectors tell me a particular tooth is from a dolphin
v. a porpoise, I have no idea what (if any) criteria is used. The
terms dolphin and porpoise have changed in meaning through time, and
formerly were actually synonyms. Many delphinid dolphins as recently
as the 1970s were referred to as porpoises. The current meaning
refers to the Phocoenidae – a half dozen species of small bodied,
short-snouted delphinoids typically found in temperate to subpolar
coastal environments around the globe. All modern phocoenids share
spatulate teeth – they almost resemble the tooth of an ankylosaur
if all the little cusps were eroded. The crowns of the teeth are
shaped like little spatulas or shovels, whereas the teeth of oceanic
dolphins (Delphinidae) are almost entirely conical (which is
primitive for most odontocete groups). True spatulate teeth
representing phocoenids are very rare in the rock record – most of
them are from the North Pacific and the west coast of South America,
dating back only 10-12 million years (possibly older in Japan,
judging from some fascinating research presented last year at SVP).
The Atlantic coastal plain has zero published records of fossil
phocoenids, and I have only seen a single spatulate tooth from the
east coast – currently in a private collection, and if I recall
correctly, from the Peace River in Florida (Plio-Pleistocene). There
are a couple of phocoenid species from Pliocene deposits in Belgium,
suggesting that we should find them in the western North Atlantic
Pliocene – but evidence so far has not been forthcoming. Further
complicating matters is that many extinct phocoenid porpoises
actually primitively retain conical teeth, indicating that only the
most modernized porpoises evolved the characeristic spatulate teeth.
Many archaic phocoenids from the Pacific retain asymmetrical
delphinid-like skulls, blurring the lines between the families. So
the take home message is this: 1) porpoise has a very specific
meaning (Phocoenidae) and 2) true porpoises (Phocoenidae) are
currently known reputably from fossil sites along the margins of the
Pacific and North Sea only – meaning if you are an east coast
fossil collector, you have probably not found a porpoise.
L. G. Barnes and R. E. Reynolds. 2009.
A new species of early Miocene allodelphinid dolphin (Cetacea,
Odontoceti, Platanistoidea) from Cajon Pass, Southern California,
U.S.A. Museum of Northern Arizona Bulletin 65:483-507
W. B. Benham. 1937. The skull and other
parts of the skeleton of Prosqualodon hamiltoni. Transactions
of the Royal Society of New Zealand 67(1):8-14
M. Churchill, M. Martinez-Cáceres, C. Muizon, J. Mnieckowski, and J. H. Geisler. 2016. The Origin of High-Frequency Hearing in Whales. Current Biology 26:1-6
M. Churchill, M. Martinez-Cáceres, C. Muizon, J. Mnieckowski, and J. H. Geisler. 2016. The Origin of High-Frequency Hearing in Whales. Current Biology 26:1-6
T. T. Flynn. 1948. Description of
Prosqualodon davidi Flynn, a fossil cetacean from Tasmania.
Transactions of the Zoological Society of London 26:153-195
R. E. Fordyce. 1991. A new look at the
fossil vertebrate record of New Zealand. Vertebrate Palaeontology
of Australasia 1191-1316
R. E. Fordyce. 1994. Waipatia
maerewhenua, New Genus and New Species, Waipatiidae, New Family,
an archaic late Oligocene dolphin (Cetacea: Odontoceti:
Platanistoidea) from New Zealand. Contributions in Marine Mammal
Paleontology Honoring Frank C. Whitmore Jr., Proceedings of the San
Diego Society of Natural History 29:147-176
R. E. Fordyce and C. D. Roberts. 2009.
Fossil Pinnipedia and Cetacea. In D. P. Gordon (ed.), New Zealand
Inventory of Biodiversity 553
M. D. Gottfried, D. J. Bohaska, and F.
C. Whitmore, Jr. 1994. Miocene cetaceans of the Chesapeake Group.
Proceedings of the San Diego Society of Natural History
29:229-238
R. Kellogg. 1931. Pelagic mammals of
the Temblor Formation of the Kern River region, California.
Proceedings of the California Academy of Science
19(12):217-397
T. Kimura and L. G. Barnes. 2016. New
Miocene fossil Allodelphinidae (Cetacea, Odontoceti, Platanistoidea)
from the North Pacific Ocean. Bulletin of the Gunma Museum of
Natural History 20:1-58
R.W.
Purdy. 1996. Paleoecology of white sharks; pp. 67-78 in A. P. Klimley
and D. G. Ainley (eds.), Great White Sharks: The Biology of
Carcharodon carcharias. Academic Press, San Diego, California.
M. R. Sanchez-Villagra, R. J. Burnham,
D.C. Campbell, R.M. Feldmann, E.S. Gaffney, R.S. Kay, R. Lozsan, R.
Purdy, and J.G.M. Thewissen. 2000. A new near-shore marine fauna and
flora from the early Neogene of northwestern Venezuela. Journal of
Paleontology 74(5):957-968
T. Yuji. 2014. A dense occurrence of teeth of fossil "mako" shark ("Isurus" hastalis: Chondrichthyes, Lamniformes) associated with a balaenopterid-whale skeleton of the late Miocene Pisco Formation, Peru, South America. Bulletin of the Gunma Museum of Natural History 187:77-86.
T. Yuji. 2014. A dense occurrence of teeth of fossil "mako" shark ("Isurus" hastalis: Chondrichthyes, Lamniformes) associated with a balaenopterid-whale skeleton of the late Miocene Pisco Formation, Peru, South America. Bulletin of the Gunma Museum of Natural History 187:77-86.
Y. Tanaka and R. E. Fordyce. 2014.
Fossil dolphin Otekaikea marplesi (Latest Oligocene, New
Zealand) expands the morphological and taxonomic diversity of
Oligocene cetaceans. PLoS One 9(9):e107972
5 comments:
Hi,
Although certainly rare, I've seen genuine megalodon teeth exceeding 7 inches by a good margin. Gordon Hubbell owns one well known at 7 3/8" and 5.5 " wide. A collector from Peru owns one slightly larger. The Black Hills tooth ( from Peru too) is comparable to Hubbell's tooth.
This is an excellent read full of important information for collectors.
I know I've had several people bring me "porpoise" teeth they've collected around here, yet I have yet to have someone show me what features they saw to make that designation. Typically the reasoning is, "____ told me it was a porpoise."
Looking forward to Bobby to continue to review topics and specimens in the US for years to come.
Perhaps when the Calvert Cliffs marine mammal chapters are printed a more thorough review of that content; including species listed and the existing unnamed specimens in various museum's collections from the Calvert Formation will get a look.
Very interested to see Bobby's opinion on that; specifically squalodonts now that Butch Dooley is no longer in the area.
Anonymous: fair enough, but those are indeed specimens in private collections.
Matt and Paul: thanks! I need to get better about posting regularly. Other than the 2016 marine mammal review, other possible upcoming topics include:
-the cetothere revolution
-some sort of a post summarizing my Ph.D. research on eomysticetids
-paleo research tips V: figure construction
-aetiocetids, baleen, and suction feeding
Post a Comment