
For other entries in this series, see:
Walrus Evolution, part 1
Walrus Evolution, part 2
Walrus Evolution, part 4
Walrus Evolution, part 5
Walrus Evolution, part 1
Walrus Evolution, part 2
Walrus Evolution, part 4
Walrus Evolution, part 5

The fossil that started it all - the weird holotype mandible of Dusignathus santacruzensis.
In the 1920’s, a rather strange pinniped fossil was collected from a cliff near the Santa Cruz wharf. In 1927, the fossil – including a fragmentary skull and a well preserved pair of mandibles – was described by preeminent marine mammal paleontologist Remington Kellogg (the same guy who described Allodesmus, Neotherium, and a ton of fossil cetaceans) as Dusignathus santacruzensis. The name Dusignathus means “jaw of the setting sun”, referring to the occurrence of this fossil on the west coast. Dusignathus santacruzensis was originally thought to represent a strange sea lion; the fragmentary skull shows that Dusignathus had a low sagittal crest, procumbent but gracile upper canines (later suggesting to other workers that the type specimen is a female), single rooted teeth with simple bulbous crowns, and a robust mandible with a procumbent canine, short lower toothrow, and an enlarged flange for the insertion of the digastric muscle. Furthermore, the mandible of Dusignathus santacruzensis is unique amongst pinnipeds in having the left and right canines nearly contacting medially, and having the left and right mandibles diverge from the chin at a relatively wide (60˚) angle. Because of its incompleteness, Dusignathus continued to be enigmatic for another 50 years until Repenning and Tedford (1977) published the pinniped bible. Thanks to the realization that other fossil pinnipeds like Neotherium and Imagotaria were early sea lion-like walruses, Repenning and Tedford (1977) identified Dusignathus as another walrus, albeit a stranger one. However, in 1962, Ed Mitchell described a partial forelimb (from the type horizon of Dusignathus) with very robust ulna and radius which he identified as an unknown walrus; this specimen was later identified by Repenning and Tedford (1977) as possibly representing Dusignathus santacruzensis. They also referred some isolated Imagotaria-like bones from the Purisima Formation to Dusignathus, and subsequent discoveries of associated postcrania from the Purisima Formation and the St. George Formation near Crescent City have confirmed the referral of these postcranial elements (more on that in the future). Repenning and Tedford (1977) erected the subfamily Dusignathinae, in which they included Dusignathus, Imagotaria, and Valenictus.
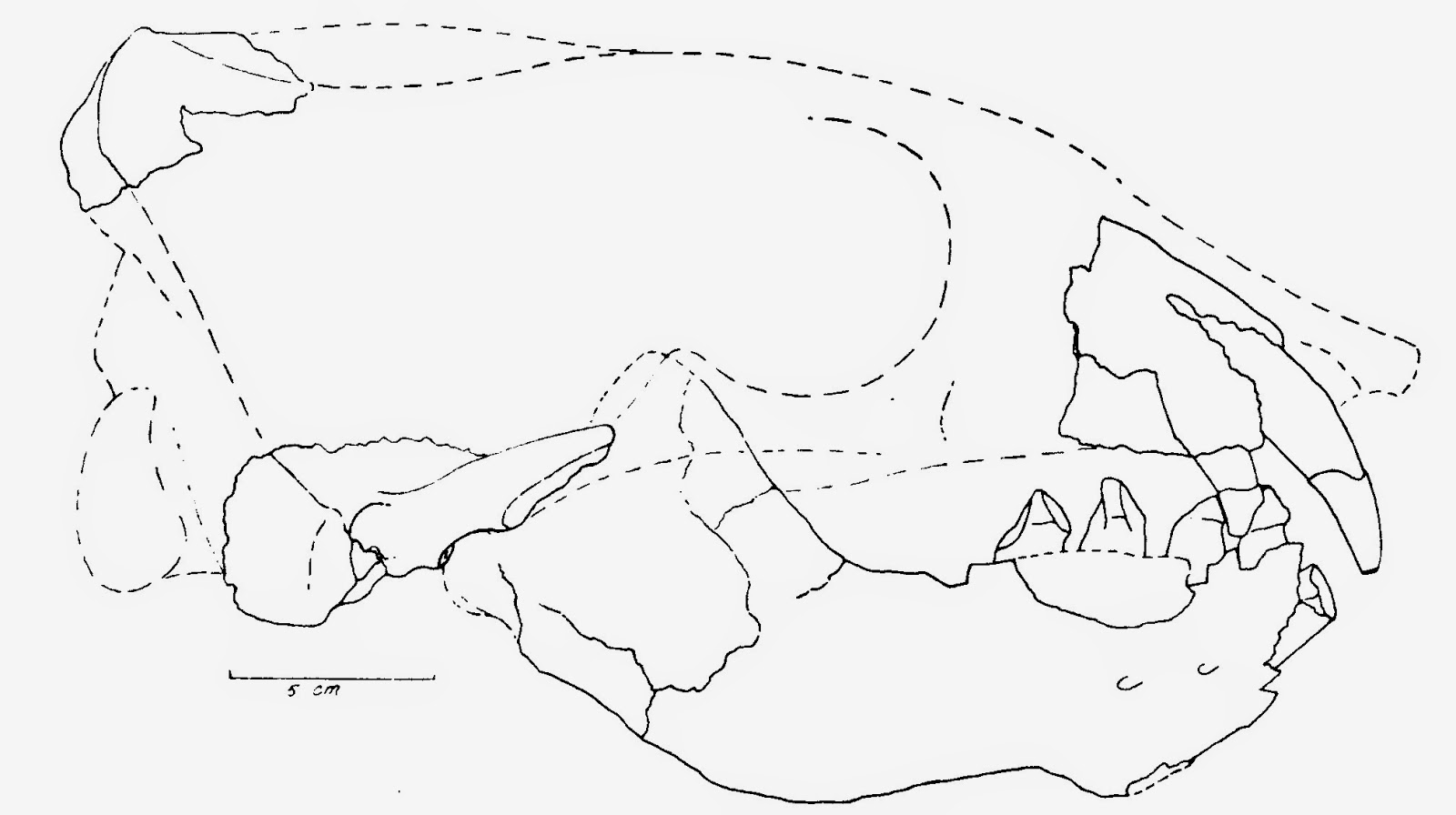
The first cranial restoration of Dusignathus santacruzensis, from Mitchell (1975). Compare this with the updated reconstruction provided below, in which I based missing skull parts and proportions on more complete material of Dusignathus seftoni.

Mitchell's (1962) walrus forelimb from the Purisima Formation at Santa Cruz, which now appears to actually represent Dusignathus santacruzensis (based on unpublished studies by yours truly).
In 1980, a gigantic walrus skeleton was discovered in exposures of the Capistrano Formation in the city of San Clemente in Orange County, California. This large walrus was named Gomphotaria pugnax in 1991 by Larry Barnes and Rodney Raschke – it roughly means “pugnacious bolt-toothed sea lion”. The type specimen of Gomphotaria includes a nearly complete, 40 cm long skull (2/3 the size of Pontolis magnus) with a large, California sea lion-like sagittal crest that would’ve given the animal a domed forehead in life, a pair of enormous but short and procumbent (e.g. nearly horizontal) tusks, single rooted and highly abraded cheek teeth, and a robust lower jaw with enlarged, procumbent lower tusks. That’s right – Gomphotaria had upper and lower tusks! In addition to the upper and lower tusks, the upper third incisor was also tusk-like and procumbent, giving Gomphotaria a third pair of smaller tusk-like teeth. Further unlike the modern walrus, Gomphotaria had a mostly flat palate – which indicates that Gomphotaria was unlikely to have been an effective suction feeder like modern Odobenus. Modern Odobenus has a highly vaulted palate which allows the tongue to act as a piston and generate tremendous oral suction. Using this piston, the modern walrus will position a clam shell in its large fleshy lips and suck the meat right out of the shell, without the shell ever entering the oral cavity. Gomphotaria, on the other hand, was probably not capable of this extreme style of suction feeding (although it must be stated that most generalized pinnipeds without arched palates, like seals and sea lions, are capable of limited suction feeding when capturing fish). Because of the rather extreme wear on the postcanine teeth and tusks of Gomphotaria, Barnes and Raschke (1991) hypothesized that it was similarly a molluskivore, but rather than sucking the meat right out of poor little clam shells – Gomphotaria simply crushed the clams to bits, shell and all. The forelimb of Gomphotaria is relatively short and robust, similar to Imagotaria, and a partial forelimb from the Purisima Formation (originally described by Mitchell, 1962) tentatively referred to Dusignathus santacruzensis by Repenning and Tedford (1977). Barnes and Raschke (1991) pointed out that several problematic walrus specimens – including Mitchell’s (1962) Purisima Fm. forelimb, and the poorly known odobenine Pliopedia pacifica (more in the next post!) ought to be reevaluated in the context of (then) newly discovered walruses like Gomphotaria.

The behemoth of a skull of the holotype specimen of Gomphotaria pugnax, photographed at LACM. The tusk is at least as big around as a beer bottle.

The holotype right forelimb of Gomphotaria, which is pathologically fused at the elbow joint.

Morgan photographing the monstrous holotype mandible at LACM in January 2012, as part of our Pelagiarctos study.
Some years after Gomphotaria was dug up in Orange County, a similar find was made further south in San Diego. A handful of specimens including an immature skull, a gigantic mandible, and a couple of humeri were discovered during a similar paleontological mitigation project. These fossils were given the name Dusignathus seftoni in a paper published by Dr. Tom Deméré in 1994, and were collected from the Pliocene San Diego Formation (4.5-2 Ma). The skull and mandible share several similarities with the somewhat older (late Miocene) Dusignathus santacruzensis, including somewhat procumbent upper and lower canines, a low sagittal crest, and a mandible with a sinuous ventral margin, medially appressed lower canines, and widely divergent mandibles. The skull of Dusignathus seftoni preserves much of the skull missing in the species from Santa Cruz, such as a relatively short and wide rostrum, a V-shaped frontal/maxillary suture, and a skull that is otherwise reminiscent of Gomphotaria and Imagotaria in general proportions. Unfortunately, the teeth of the type skull and referred mandible are poorly preserved, precluding Deméré (1994) from making any specific interpretations about feeding adaptations of Dusignathus seftoni, although general cranial similarities with Imagotaria suggested it may have been piscivorous (Demere, 1994). However, subsequently discovered skulls and mandibles with complete canines demonstrate that Dusignathus seftoni had upper and lower tusks just like Gomphotaria – albeit somewhat less procumbent and shorter. Furthermore, these teeth are highly worn, also similar with Gomphotaria. In my opinion, the dentition and toothwear of these two dusignathines are similar enough to confer a similar shared feeding ecology to both species. Extreme toothwear in Dusignathus seftoni therefore may also suggest benthic molluskivory like Gomphotaria. Additional specimens of Dusignathus seftoni now show that skull size of adult males ranged up to 40 cm (just slightly smaller than Gomphotaria and 2/3 the size of Pontolis) and that the species was (unsurprisingly) sexually dimorphic. The humerus of Dusignathus seftoni is relatively similar to Imagotaria but slightly more gracile than the isolated humerus referred to Dusignathus santacruzensis by Repenning and Tedford (1977).

The beautifully preserved holotype subadult male skull of Dusignathus seftoni in anterior view, photographed at the San Diego Natural History Museum in January 2012.

An even more beautifully preserved referred skull of Dusignathus seftoni, which is currently on display in the fossil gallery at the San Diego Natural History Museum.
As discussed in the previous post, Deméré (1994) considered Pontolis
magnus to represent a dusignathine walrus (see part 2 for a thorough
discussion of Pontolis). This issue brings up the question of
dusignathine phylogeny. Prior to the discovery of Dusignathus seftoni
and Gomphotaria, Dusignathus was considered by Kellogg (1927) to
preserve sea lion and walrus-like features; later, the Dusignathinae was
considered by Mitchell (1968, 1975) to only include Dusignathus, and
although previously recognizing walrus-like features, interpreted Dusignathus
to be more closely related to desmatophocid seals. Repenning and Tedford (1977)
later included within the subfamily Imagotaria downsi (based on
similarly sized upper and lower canines) as well as the poorly known walruses Valenictus
imperialensis and Pliopedia (based on shared robust proportions of
forelimb bones). As it turns out, Valenictus and Pliopedia were
later recognized to represent tusked walruses (Odobenines; Deméré, 1994).

The early phylogenetic hypothesis of otarioid phylogeny from Mitchell (1975).

One of the first computer-aided cladistic analyses of walrus phylogeny - from Deméré (1994).
Early studies presented phylogenetic hypotheses that were not rigorously or systematically conducted. Prior to the advent of computer-aided cladistics, the majority of phylogenetic hypotheses were formulated in a subjective manner and cladograms were hand-drawn – often with all of the intellectual baggage from the author’s own preconceived notions of relationships intact and unchecked. Fortunately, those dark days are now (mostly) over, and rather than risk making dubiously informed claims of phylogeny based on subjective interpretations of a handful of anatomical characters (e.g. using primarily earbone characters over features of the rostrum and palate for whales and dolphins). Now that we have computers, we can statistically evaluate phylogenetic hypotheses using hundreds (or more) of anatomical characters (my own dissertation on baleen whales features a large dataset with over 300 morphological characters) scored for many species. The first computer-aided analyses of walrus phylogeny were published by Tom Deméré (1994) and Naoki Kohno (1994). These early studies used relatively small datasets (53 and 25, respectively); a later study by Deméré and Berta (2001) on Proneotherium featured only 24 characters. The cladistic analysis of Deméré (1994) confirmed the monophyly of the dusignathine subfamily, and also – using newly recognized USNM specimens that Doug Emlong himself collected in the 1960’s and 1970’s from the Empire Formation – recovered the giant walrus Pontolis magnus as another dusignathine. Kohno (2006) pointed out that the limited dataset used by Deméré and Berta (2001) did not necessarily result in a more accurate phylogeny, and assembled a new cladistic matrix for walrus phylogeny using 60 characters. When Morgan and I started work on Pelagiarctos back in 2012, we used Naoki Kohno’s 2006 matrix as a starting point, and added a ton of mandibular characters so that we could place Pelagiarctos – still only known from a mandible – into a phylogenetic context (our matrix had 90 characters). Interestingly, my labmate Yoshi Tanaka (and his master’s adviser, Dr. Kohno) is working on a new fossil “imagotariine” from the same locality as Pseudotaria muramotoi – and has used our modified matrix and added a new character and modified several others. So, the walrus matrix has been batted around full circle now (or, soon will be).

An improved, larger cladistic analysis by Kohno (2006).
So what happened when additional characters were added? Dusignathines remained monophyletic but Pontolis dropped out and appeared as a sister species to Imagotaria in Kohno’s (2006) study; this is perhaps unsurprising, since Pontolis has an Imagotaria-like basicranium and fossils of the two have been frequently mistaken for one another since the 1940’s (e.g. Lyon, 1941; Mitchell, 1968). Our own study found a compromise of sorts between Pontolis-as-a-dusignathine and Pontolis-as-sister-to-Imagotaria: Pontolis in our study was recovered between the Pelagiarctos-Imagotaria clade and the Dusignathinae (Boessenecker and Churchill, 2013). Furthermore, support for the Dusignathinae decreased a bit. Some of the features which Deméré (1994) noted as dusignathine synapomorphies present in Pontolis include a V-shaped frontomaxillary suture – however, this condition characterizes some fossil odobenines as well; a tusk-like lower canine may genuinely support such a relationship, but a sinuous ventral margin of the mandible (as in Dusignathus spp.) is present in some specimens of Imagotaria.

An even larger study, from our own paper on Pelagiarctos (Boessenecker and Churchill 2013).

Skull diversity amongst the dusignathines, with Pontolis included as it may yet represent a dusignathine, and Neotherium for comparison. These bigger, freakier walruses make Neotherium look generalized, small, and sort of adorable in comparison.
Lastly, to follow up this rather dry phylogenetic talk – what is the evolutionary significance of the dusignathines? Why are they my favorite of all walruses? To sum that second question up, in my mind they are a relatively strange and highly derived group with a strange dentition (lower tusks, and procumbent upper tusks!), in some cases large sagittal crests paralleling some sea lions, and strange postcranial features such as unusually robust, shortened forelimbs. They are the least studied and most neglected group of walruses by far, and also geographically limited: the entirety of the dusignathine fossil record is from California, Oregon, and Baja California. These pinnipeds evolved during a time of relative faunal isolation: warm equatorial waters barred dispersal to the southern hemisphere or dispersal through the Panama Seaway. The Bering strait had not yet opened until the latest Pliocene, barring dispersal to the Atlantic – and for some strange reason, dusignathines never made it to the western North Pacific (as evidenced by their conspicuous absence in Japanese marine mammal fossil assemblages), unlike virtually every other North Pacific pinniped group. This is not to say that I am any less passionate about odobenines, which certainly had some serious weirdos in their own right during the Pliocene – but these dusignathines evolved somewhat in parallel but in a different direction, and represent an extinct group of highly derived walruses most people are unfamiliar with but dominated over 1000 miles of the western coastline of North America until only 2 million years ago. In a way, dusignathines can be viewed as another mini radiation of specialized walruses that evolved in parallel with the tusked odobenines, snuffed out during the end-Pliocene marine mammal extinction for some unknown cause. What questions and research has yet to be done? For starters, at least two new genera of dusignathines (...or possible imagotariines) have yet to be described, and referred material of Pontolis magnus is desperately needing description (this may be the next joint project Morgan and I tackle). The postcranial skeleton of Gomphotaria begs a complete description – and enough new material of Dusignathus santacruzensis now exists to make sense of a wide variety of seemingly unrelated postcranial bits scattered between various collections and publications. Dusignathus seftoni is now the most completely known dusignathine, now known from three or four skulls, almost as many mandibles, a couple of partial skeletons, and a host of postcrania. We still don’t really know what these guys were feeding upon (at least, no more than the idle speculation provided above), and toothwear, morphometric studies, and isotopic analysis could resolve that question. Also, why were their foreflippers so short and stubby? As you can see, there’s a lot left to do.
Next up: the fossil record and evolutionary biogeography of
true “tusked” walruses (Odobeninae).
References
Barnes, L.G. and R. E. Raschke. 1991. Gomphotaria pugnax, a new genus and species of late Miocene dusignathine otariid pinniped (Mammalia: Carnivora) from California. Contributions in Science 426:1-16
Barnes, L.G. and R. E. Raschke. 1991. Gomphotaria pugnax, a new genus and species of late Miocene dusignathine otariid pinniped (Mammalia: Carnivora) from California. Contributions in Science 426:1-16
Boessenecker, R.W., and M. Churchill. 2013. A Reevaluation
of the Morphology, Paleoecology, and Phylogenetic Relationships of the
Enigmatic Walrus Pelagiarctos. PLoS One 8(1):e5411.
Deméré, T.A. 1994. The Family Odobenidae: A phylogenetic analysis of fossil and living taxa. Proceedings of the San Diego Society of Natural History 29:99-123.
Deméré, T.A. 1994. Two new species of fossil walruses
(Pinnipedia: Odobenidae) from the Upper Pliocene San Diego Formation, California.
Proceedings of the San Diego Society of Natural History 29:77-98.
Kellogg, R. 1927. Fossil Pinnipeds from California.
Contributions to Palaeontology from the Carnegie Institution of Washington
27-37.
Kohno, N. 1994. A new miocene pinniped in the genus Prototaria (Carnivora: Odobenidae) from the Moniwa Formation, Miyagi, Japan. Journal of Vertebrate Paleontology 14(3):414-426.
Kohno, N. 1994. A new miocene pinniped in the genus Prototaria (Carnivora: Odobenidae) from the Moniwa Formation, Miyagi, Japan. Journal of Vertebrate Paleontology 14(3):414-426.
Kohno, N. 2006. A new Miocene odobenid (Mammalia: Carnivora)
from Hokkaido, Japan,
and its implications for odobenid phylogeny. Journal of Vertebrate Paleontology
26(2):411-421.
Lyon, G. M. 1941. A Miocene sea lion from Lomita,
California. University
of California Publications in
Zoology 47:23-41.
Mitchell, E.D. 1962. A walrus and a sea lion from the
Pliocene Purisima Formation at Santa Cruz, California:
with remarks on the type locality and geologic age of the sea lion Dusignathus
santacruzensis Kellogg. Los Angeles
County Museum
Contributions in Science 56:1-24.
Mitchell, E.D. 1968. The Mio-Pliocene pinniped Imagotaria.
Journal of the Fisheries Research Board of Canada
25(9):1843-1900.
Mitchell, E.D. 1975. Parallelism and convergence in the
evolution of the Otariidae and Phocidae. Rapports et Proces-verbaux des
Reunions 169:12-26.
Repenning, C.A.
and R. H. Tedford. 1977. Otarioid seals of the Neogene. Geological Survey
Professional Paper 992:1-93.
No comments:
Post a Comment