
Merry Christmas/Happy Hannukah/Chrimbus/Festivus! Welcome to the third annual review of
advances in marine mammal paleontology! There were over 50 new publications in
marine mammal paleontology this year, and this took quite a while to work up;
about 1/3 of the papers I didn’t even really get a chance to read until this
winter (er, summer). As usual, this list is supposed to be comprehensive, so if
I have missed something, please let me know – hopefully I’ll have time to
update it. I may not, because ultimately my thesis is more important. As in the
past two years, I include papers that have been published since January 2014 –
however, for papers that came out online in 2013 and were only included in
published journal issues in 2014 – these will remain in last year’s post (and
the same goes for some of the following studies, some of which will almost
certainly have permanent 2015 citation dates despite online publication in late
2014). There are three or four I still have yet to add, so stay posted and check again in a few days.
It does bug me when I see new papers that
have left out citations to important new research; I try to devote some time to
reading new publications as they come out, and hopefully with these posts
statements like “well I didn’t really know about such and such article, I don’t
know how to use google” or “I’ve literally been living under a rock” will no
longer be decent excuses. Anyway, I hope this huge review will be of service for
other marine paleomammalogists!

This paper formed one of the core chapters
of former labmate Gabriel Aguirre-Fernandez’ doctoral thesis, and reports a
well-preserved and beautifully prepared odontocete skull from the lower Miocene
Kaipuke Siltstone in northwest Nelson, South
Island, NZ. This specimen was for years
thought to be the earliest record of kentriodontids (but keep reading). The
skull is rather small, perhaps similar in size to a Hector’s dolphin or harbor
porpoise. It appears to have had a polydont single rooted dentition and a
subtriangular, acutely pointed rostrum and convex palate – overall not terribly
different from a dwarfed Waipatia aside from the tooth rooting.
Cladistic analysis indicated rather strongly that this is no kentriodontid, and
is in fact a stem odontocete of uncertain position; it’s more derived than
xenorophids (see Geisler et al., below, for more on xenorophids), but more
primitive than beaked whales and sperm whales. They named the new dolphin Papahu
taitapu, meaning dolphin from Te Tai Tapu (the area of Nelson where it was
collected from) in Māori.

One of the most profound and strangest
discoveries in marine mammal paleontology is the discovery of aquatic adapted
sloths from the Miocene and Pliocene of Peru (and now Chile as
well). These were first named Thalassocnus in 1995, and in the following
decade a number of additional species and a ton of skeletons were reported and
described. Common questions I get about these from colleagues in
non-mammalogical fields of paleontology are “How do we know they’re actually
aquatic and not just terrestrial sloths that floated out to sea?”. It’s an
excellent question that can be addressed taphonomically and adaptationally.
Taphonomically speaking, these skeletons have only been found in marine
sediments and none yet from terrestrial deposits – which have other types of
sloths. Further, apparently much of the coastline of Peru
during the late Neogene was inhospitable desert. These sloths also have
modified limb proportions and large muscle attachment areas on caudal vertebrae
for undulation of the tail (if I recall correctly; it’s been almost ten years
since I last read the original studies). Another approach – taken by the new
article by Eli Amson - is paleohistology. Most tetrapods that return to the
water go through a period where postcranial bones become denser, as a buoyancy
adaptation thought to offset positively buoyant air-filled lungs during shallow
diving. This pattern of increased bone density (pachyostosis – increased
cortical bone thickness; osteosclerosis – decreased medullary cavity volume;
and pachyosteosclerosis – combination of both) characterizes the early
evolution of most marine tetrapod groups, including (but not limited to) ichthyosaurs,
mosasaurs, pachypleurosaurs, placodonts, most flightless diving birds
(penguins, mancalline auks), pinnipeds, sirenians, cetaceans, desmostylians,
and sea otters. These patterns of bone thickness are to a degree obvious
externally when comparing with terrestrial ancestors, particularly with modern
specimens where differences in bone mass and density are easily felt during
handling. In fact, using Archimedes principle is an easy way to determine bulk
bone density for modern specimens – but for fossils, there is always an unknown
mass of sediment and cement inside, and the vagaries of diagenesis otherwise
make this approach impossible. Instead, we can either chop up bones and look at
them under a microscope – or use CT scanning to get a cross section. In either
case a software program called Bone Profiler can be used to quantitatively
compare cortical bone thickness and a dimensionless compactness metric across
taxa. Amson et al. did just this with ribs, femora, and tibiae of three species
of Thalassocnus (T. antiquus, T. natans, T. littoralis,
and T. carolomartini) and three outgroups (an anteater, an extant sloth,
and the extinct ground sloth Hapalops) and found that bone density does
in fact increase from the basal end of the tree to the end. The three species
of Thalassocnus are stratigraphically separated and perhaps thought to
represent cross-sections of a lineage through time, and interestingly (and
predictably) bone thickness increases in younger species. As an aside, some
groups that exhale and let their lungs collapse – an adaptation for deep diving
– have actually gone the other way around, and evolved light porous bone
(osteoporotic – yes, the very same condition osteoporosis that
disproportionably affects women) to offset the loss of buoyant air-filled
lungs, and includes derived ichthyosaurs, most modern cetaceans (particularly
odontocetes), elephant seals, and a single desmostylian (Desmostylus).
I’ve taken a bunch of thin sections of eomysticetid ribs, and have tinkered
around with converting images of my thin sections for analysis in Bone Profiler
(needs more tinkering still).

Egypt and Pakistan are perhaps the two countries most famous for producing spectacular
fossils of archaeocete whales that have provided us with the skeletal evidence
demonstrating the famous land to sea transition ("feet to flippers")
that is now a poster child for vertebrate macroevolution. Although often
outshined by Pakistan, India has produced a number of important fossils as well, including
pakicetids, remingtonocetids, and protocetids; unsurprisingly, most of these
fossils come from Kutch, the westernmost part of India
that is fairly close to Karachi, the capital of Pakistan.
This new study by Bajpai and Thewissen reports several new protocetid specimens
from India. Some of these include a new partial mandible and a maxilla
fragment with teeth, the first for the earlier described protocetid Indocetus
rahmani. They also describe two new genera of protocetids; the first, Kharodacetus
sahnii, is represented by a well-preserved anterior skull and partial
mandibles with well-preserved teeth, and a few referred cranial specimens. Kharodacetus
is one of the largest known protocetids worldwide, and is similar in size to
the basilosaurid Zygorhiza - heralding the transition from medium-sized
protocetids to the larger body sizes attained by later basilosaurids. The
second new genus is based upon a nearly complete but poorly preserved skull,
and is named Dhedacetus hyaeni (after the hyenas that live at the
type locality!). Dhedacetus also has a referred vertebral column, and
this column indicates it had a tail with robust musculature, indicating
tail-based propulsion (rather than hindlimb propulsion as proposed for Georgiacetus
by Mark Uhen a few years ago). Bajpai and Thewissen further point to Maiacetus
(one of the only completely known protocetids) and suggest it may have actually
had a caudal peduncle and fluke (although see Uhen 2014 on Natchitochia,
below).

The early Miocene squalodelphinid dolphin Notocetus
vanbenedeni was originally reported from Argentina.
Notocetus vanbenedeni is a long-snouted archaic dolphin, and the
squalodelphinids are a group of platanistoids – a formerly diverse group of
odontocetes more closely related to the modern ganges river dolphin, Platanista.
Another platanistoid – Otekaikea marplesi – was formerly placed in Notocetus
by my adviser Ewan Fordyce in the 1990’s, but a redescription of that
odontocete indicates it’s more closely related to Waipatia (see below:
Tanaka and Fordyce). Other squalodelphinids include Squalodelphis (Italy),
and most importantly, Huaridelphis (see Lambert et al., below). Huaridelphis
was also collected from the Chilcatay Formation, indicating that two
squalodelphinids were likely sympatric. Notocetus vanbenedeni is now
known from both the west and east coast of South
America during the early Miocene.

As discussed elsewhere below, there has
been an explosion of research on the morphology, phylogeny, and feeding ecology
of true cetotheres, the Cetotheriidae sensu stricto. One such new
cetothere, one that is obviously closely related to my favorite cetacean, the
dwarf baleen whale Herpetocetus, is reported in this new paper by
Michelangelo Bisconti and is named Herentalia nigra. It’s based on Herpetocetus-like
braincase from the upper Miocene of Belgium, and differs principally from Herpetocetus
in only a few features – namely shorter ascending maxillae, longer exposure of
the parietal at the “vertex”, a weird periotic, a squamosal cleft, and is much
larger in absolute size. It is nearly identical to Piscobalaena nana
from coeval strata in Peru, differing only in its larger size and presence of a squamosal
cleft – which makes me wonder whether it might be a species of Piscobalaena.
This study also criticizes the recent hypothesis that the pygmy right whale is
an extant cetothere, even with a section of the discussion titled “Is Caperea
marginata a cetotheriid sensu stricto?”. Bisconti brings up many
points of disagreement on coding of various characters in the cladistic matrix,
focusing on characters and character state definitions identified as supporting
the relationship. Bisconti’s cladistic study instead supports the traditional
view of balaenoid monophyly – supported by virtually all analyses aside from
those by former labmate Felix Marx. I have my own reservations, but those will
have to wait until my own enormous mysticete matrix from my doctoral thesis
gets published (currently in review).

This paper was the first from my doctoral
thesis to be published, and describes new (and old) eomysticetid material from
the south island of NZ. The fragmentary skull and earbones of “Mauicetus” waitakiensis
were never really properly described or figured, and more recent discoveries of
the type species of Mauicetus (Mauicetus parki) reveal that “Mauicetus”
waitakiensis belongs to a more primitive group of mysticetes. Its
incompleteness, however, hampered any attempts to interpret it. A new skull
with other associated bits (tympanoperiotic, mandible, vertebra, ribs) shared
unique features of the posterior braincase and tympanic bulla of “Mauicetus”
waitakiensis, convincingly linking the two together. So, we named the
new genus Tohoraata raekohao for the more recently collected skull, and
recombined the older species as Tohoraata waitakiensis. Tohoraata is
Māori for “Dawn whale”, in parallel with the closely related “Eomysticetus”,
which means the same thing. Cladistic analyses including Tohoraata will
be published in a subsequent paper derived from my thesis research.

Bone-eating worms – Osedax – infest
skeletons at modern whale whale falls on the deep sea floor, boring little
holes into bone and digesting/dissolving bony tissues with the help of
symbiotic bacteria. This activity leaves known traces in modern whale bones,
and fossil whale bones from the Oligocene of Washington (USA) and Pliocene of
Italy have already been found with Osedax traces. This study presents
the first record of bone eating worm fossil traces from the Southern Hemisphere
– which in and of itself is noteworthy, but not exactly groundbreaking. Rather,
the traces we found on an Oligocene eomysticetid skull and mandibles are
cross-cut with tooth marks from a shark or bony fish. Because Osedax generally
colonize skeletonized bones (e.g. defleshed), these tooth marks are not
attributable to scavenging of remnant soft tissues but instead reflect feeding
upon the soft tissues of the worms themselves, which do leave a thin protective
barrier of outermost bone – but generally too thin to dissuade vertebrate
predators. Interestingly, modern ratfish and crabs have been observed ripping Osedax
out of bones at whale falls.

Taphonomy is the study of fossil
preservation, essentially including all processes that affect a dead organism
from death until burial. It’s easy to study the actualistic taphonomy of
terrestrial organisms, since those are environments accessible to us: we can
watch a dead animal decompose our backyard, or chuck bones into a stream (or
flume) and watch them get sorted. We’re less lucky with marine vertebrates,
however; actualistic studies effectively include studies of carcasses and bones
along shorelines, or studies of deep sea whale falls. These mark important
contributions, but tell us virtually nothing about what happens in between on
the continental shelf – a setting reflecting virtually all Cenozoic marine
vertebrate assemblages. Instead, we can turn to the rock record for a
historical perspective. For my master’s research I was interested in
broad-scale patterns in preservation of vertebrate skeletal material in shelf
sediments. Most shelf deposits don’t preserve bones and teeth in numerous
settings, and as such most prior taphonomic studies focused on single bonebed,
skeleton, or lithofacies. The Purisima Formation in California is
unique as it preserves abundant marine vertebrates in outer, middle, and inner
shelf settings, permitting study of across-shelf changes in preservation. I
collected data from over a thousand specimens (many of which I excavated
myself) including abrasion, articulation, phosphatization, fragmentation, and
polish. As it turns out, the intensity and style of preservation changed
remarkably between environments, and was more or less correlated with inferred
depositional energy; higher energy settings nearer shore had lower rates of
articulation but higher rates of fragmentation, abrasion, phosphatization, and
polish. Even more extreme taphonomic damage characterized skeletal material
from bonebeds, which were formed largely by a combination of seafloor erosion
during transgressive episodes followed by periods of nondeposition.
Preservation also varied between taxonomic groups and tissue types – regular
bones were somewhat average, with teeth more durable, and calcified cartilage
less so. Bird bones, perhaps owing to their lower mass, were less
taphonomically modified; cetaceans had some of the highest rates of damage,
particularly fragmentation (perhaps according to osteoporosis). These
differences indicate a potential for bias regarding differential preservation,
and suggests a significant taphonomic overprint.

This paper was a long time in the making,
and had its roots in a comprehensive phylogenetic analysis of pinnipeds that
Morgan and I started putting together back in 2008 or 2009. We presented on it
in 2010, and quickly realized that the way forward was to break up the analysis
by family. Our first stab was the phylogeny presented in the Pelagiarctos
article, where we added a few new mandibulodental characters for walrus
phylogeny. Pinnipeds are certainly challenging to work on; historically there
has been a bit of debate about which characters to use, and there has also been
some rather shocking choices in character selection, and some parts of the
pinniped skeleton have been totally neglected. Anyone who has read this blog
before knows that pinniped phylogeny is fairly contentious, and perhaps not as
politely discussed as various disagreements in cetacean phylogeny, for example.
Disagreements over whether pinnipeds are monophyletic still exist
despite diphyletic hypotheses being thoroughly discarded by molecular studies decades
ago, and disagreements over the sister taxon of walruses and desmatophocids
(among others) still rage. While we will be revisiting odobenid phylogeny with
a far larger and more comprehensive analysis in the future (with spectacular
new fossils… yay!), our second foray on our long-term pinniped cladistics
‘campaign’ focused on my other favorite group of pinnipeds, the otariids – fur
seals and sea lions. Unknowns in otariid phylogeny include 1) are fur seals and
sea lions reciprocally monophyletic (e.g. Arctocephalinae and Otariinae), and
2) when did otariids make it to the southern hemisphere? We set out to clarify
these questions using morphological and molecular data. Fur seal monophyly had
been proposed by an earlier study using fewer characters, but has been
challenged by most (if not all) molecular studies which suggest that
arctocephalines are paraphyletic, if not polyphyletic. Our results confirm (for
the first time using morphological data) fur seal paraphyly, but in general
there is a fair amount of variation in the trees from different types of
analyses and search protocols. This is not quite our fault, but likely relates
to problems with fur seals and sea lions: they all look the same, resulting in
relatively few characters distinguishing between different otariids.
Furthermore, there is extreme variation amongst these characters, further
obscuring phylogenetic signal. Regardless, we did the best we could, and
produced the best analysis to date (and you can see all of our data on
morphobank!). Using updated dates of fossils from the southern hemisphere, our
results permit reconstruction of the southern hemisphere dispersal of otariids
to the latest Miocene and along the east Pacific margin, during a period of
unusually cool equatorial surface temperatures in the eastern Pacific. What do
we need to further the study of otariid evolution? More fossils and more
characters, of course! Anyway, next up in the pinniped phylo campaign is a
serious reappraisal of odobenid phylogeny (my focus), and an even more critical
reassessment of phocid phylogeny (Morgan’s focus).

Modern pinnipeds consume a variety of prey
items, including fish, krill, bivalves, cephalopods, and even birds and
occasionally other pinnipeds. Different feeding behaviors are utilized
including regular raptorial feeding (often accompanied with suction), suction
feeding, filter feeding, and macrophagy (e.g. ripping large prey items into
smaller pieces for swallowing). Such a diversity in feeding was likely present
in fossil pinnipeds. How do we tell how extinct pinnipeds ate? We can take a
stab from the “ass end”, so to speak – see if there preserved gut contents (or,
coprolites) as often are for marine mammals. Well, there are only two published
cases of pinnipeds with gut contents: one is the phocine Kawas from Argentina,
and another is a Pliocene phocid from Waihi Beach in Taranaki, New Zealand, in a private collection but “published” in an abstract by Joe
McKee. Kawas tells us that (unsurprisingly) it ate fish (in other news,
the sky is blue). Rather than using the end-product, we can also attack it from
the business end – seeing if differences in feeding morphology can be used to
predict feeding ecology, by analogy with observed feeding adaptations in modern
pinnipeds. To tackle questions like this, Morgan Churchill and colleagues used
discriminant function analysis of tooth spacing measurements for modern and
fossil pinnipeds. Earlier studies have found that only a rough correlation
between feeding morphology and ecology exists, as most pinnipeds are dietary
generalists – a finding supported by this study, which is the most thoroughly
quantitative on the subject to date. Pinnipeds that either filtered prey items
from the water column or ripped prey to pieces before swallowing generally have
larger teeth and smaller gaps between them than pinnipeds which just swallow
prey items whole. As opposed to earlier studies which identified four different
strategies – pierce, suction, filter, and grip-and-tear feeders, this study
found support for three basic groups: sieve feeders like Lobodon and Hydrurga,
large-toothed raptorial feeders, and small-toothed raptorial feeders. Mirounga
belongs to the latter category, likely reflecting tooth reduction and
specialization for suction feeding – although critically, the bearded seal (Erignathus)
has been observed suction feeding and plotted in this study with other phocine
seals, and further, the crabeater and leopard seals (Lobodon and Hydrurga)
were not predicted to be suction feeders – effectively, this not only attests
to the limited morphological adaptations for different feeding styles but also
highlights the behavioral plasticity of pinnipeds. Most critically, and why
this study is on this list as it is otherwise largely neontological in scope –
is that it reconstructed the likely feeding ecology of the fossil pinnipeds Enaliarctos
and Desmatophoca. Both extinct taxa plotted close to modern otariids,
suggesting that they were piscivores that used pierce/raptorial feeding
behavior. Part of Morgan’s original question was to figure out whether or not Enaliarctos
(the earliest known pinniped) – which primitively retains carnassial teeth –
was able to feed on fish and swallow them whole, or whether it needed to return
to shore to chew its food like its terrestrial ancestors.

Cope's rule (advanced by none other than Edward Drinker Cope) is the hypothesis that body size tends to increase through time within verebrate lineages (I'm not exactly sure if it was proposed to apply to invertebrates). This idea sought to explain trends toward gigantism, followed by extinction of a lineage, niche replacement, and trend toward gigantism in the replacing taxon. Cope was inevitaby inspired by gigantic fossils of sauropod dinosaurs, but also worked a fair amount on fossil baleen whales - which are quite a bit larger today than the majority of most fossils. Cetaceans are the obvious group to study given the amazing diversity of sizes and gigantic modern sizes - but their fossil record is a bit problematic to work with in some ways. My buddy Morgan Churchill published another recent article (not included here because it is neontologic rather than paleontologic) proposing equations to predic body size from craniomandibular measurements as part of his Ph.D. - and these were applied to fossil pinnipeds in order to examine trends in body size through time. Cope's rule - defined as consistent trends towards larger size within a lineage - is present at a very rough level; the earliest pinnipeds are generally quite small, and modern pinnipeds can be quite large. However, rather than showing a universal increase in body size amongst various lineages, body size tends to diversify through time - in other words, maximum body size increases (cf. walrus, elephant seals), but minimum body size does not (cf. baikal seals, townsend fur seal) - which appears to reflect passive radiation into different niches.

-and-
Note: I’m bundling these two papers
together since they tell the same story in differing levels of detail. Several
species of sea lions are alive today, including the northern hemisphere
California (Zalophus californianus) and Steller’s (Eumetopias
jubatus) sea lions, and southern hemisphere species such as the South
American (Otaria byronia), Australian (Neophoca cinerea),
and New Zealand sea lion (Phocarctos hookeri). I’ve seen many of
the latter while living down under, and they are mean: Zalophus back
home in California aren’t very bold and often will return to the water if you approach
them, but I’ve seen bull NZ sea lions charge idiot tourists with cameras.
Although easy to see along nearby places like the Otago Peninsula,
they don’t actually breed on the mainland – something that has puzzled
biologists for quite some time. They breed at offshore islands, like the Subantarctic
islands for example, and simply haul out on the mainland – but never stay very
long. This is puzzling, as Holocene subfossils of juveniles reported by
paleornithologist Trevor Worthy demonstrate that in prehistory they bred
onshore. These new studies by Collins et al. report that subfossil Phocarctos
from mainland New Zealand are genetically distinct from the modern
population using analysis of ancient DNA, and bred on the mainland until
becoming extinct. It’s unclear what this means as far as taxonomy is concerned
– if anything, it might mean it’s distinct only at the subspecies level. Prior
to Māori arrival, specimens that are molecularly related to the modern form are
found only in sites on the Subantarctic islands. These studies demonstrate the
extirpation of an entire population of mainland breeding sea lions (fossils of
which indicate it inhabited all of the mainland aside from the northwestern
coast of the South Island) shortly after Māori arrival (indeed, many of the
specimens sampled are from middens) and the area reinhabited by the
subantarctic population of Phocarctos, but retaining the ancestral
subantarctic breeding behavior. Interestingly, this parallels the mainland
extinction of the Waitaha penguin; within a century the areas formerly occupied
by the Waitaha penguin (Megadyptes waitaha) were recolonized by the
extant population of closely related Yellow-eyed penguins (Megadyptes
antipodes). As an aside, the second author on these papers is friend and
fellow Dunedinite Nic Rawlence.

Modern porpoises (Phocoenidae) are
relatively small bodied dolphins that generally lack “beaks” – they inhabit
much of the North and South Pacific, and North and South Atlantic. Extinct porpoises
have similar distribution, but are most abundantly known from the North Pacific
– and a North Pacific origin for the clade has been implicated. A late Miocene
dispersal to the eastern south Pacific is evident thanks to fossils like Australithax
from Peru; a single Pliocene periotic from offshore New Zealand indicates the timing of dispersal to the western South Pacific. The
North Atlantic is another story; phocoenids are totally absent from Pliocene rocks
in the eastern USA. In 2008, an archaic phocoenid, Septemtriocetus, was
reported from the “middle” Pliocene of Belgium. By this point in time the Panama
seaway was already closed to trans-oceanic dispersal, but the Bering strait had opened –
indicating that the ancestor of Septemtriocetus likely invaded the North Atlantic via the Arctic Ocean (which was not yet
glaciated). A new phocoenid, named Brabocetus gigasei, is reported in
this new study from the lower Pliocene Kattendijk Formation of Belgium –
somewhat older than Septemtriocetus, indicating an older dispersal to
the Atlantic. Brabocetus has an asymmetrical skull, and a well-preserved
braincase – effectively resembling a modern Phocoena but with an
asymmetrical facial region and a vertex – similar in many regards to Septemtriocetus,
except having a wider antorbital notch. The occurrence of these two porpoises
in the North Sea Pliocene possibly suggests two dispersals to the North Atlantic or perhaps in
situ speciation; regardless, the authors indicate that the harbor porpoise
likely represents an independent, late Pleistocene dispersal from the North
Pacific.

In 1741, German naturalist Georg Willhem
Steller set foot on Alaska for a few hours and became the first European to visit Alaska. This was
but a brief stop for the Second Kamchatka Expedition, later known as the Great
Northern Expedition - one of the largest naval expeditions, commissioned by
Emperor Peter the Great of the Russian Empire to explore the eastern reaches of
Siberia
(recently incorporated into the empire). On the return from the eastern North
Pacific, the crew succumbed to bouts of scurvy, leaving only a dozen able
bodied sailors on board, and in fall 1741 the remaining ship, the St. Peter,
was shipwrecked on a small island in the western North Pacific. The leader of
the expedition, Vitus Bering, died on December 8, and the crew was largely
marooned on the island - later named Bering Island.
Steller soon discovered bizarre, gigantic marine mammals around the island,
which resembled a manatee but had cetacean-like caudal flukes and much larger -
up to 8-9 meters in length - and fed upon kelp. It would later be named
Steller's Sea Cow (Hydrodamalis gigas) in his honor. It was very
buoyant, and part of the sea cow's back was always exposed above the water line,
and was otherwise a very slow swimmer unafraid of humans. They could be killed
simply by using grappling hooks and dragging them ashore - and worse, they
appeared to have strong individual bonds, and would attempt to come to the aid
of those being killed - which gave hunters an easy next target. The crew
eventually built a smaller boat some time after the wreck of the St. Peter was
destroyed by foul weather - and they successfully sailed back to Russian
settlements in Kamchatka. News of gigantic, easily killed marine mammals spread, and over the
next few decades many crews stopped by Bering Island to take sea cows as their
fat was excellent for burning (apparently smokeless) and served well as a
butter substitute. By 1768, less than 30 years later, all that remained were
bones. Conventional thought in sirenian biology considers this to be the last
holdout of the sea cow in the Pacific, although Pleistocene and Pliocene
fossils indicate that Hydrodamalis formerly lived as far south as Japan and
Baja California. The new study by Crerar et al. reports new bones of Hydrodamalis
from St. Lawrence Island, also in the Bering sea but further north and east
- which date to the Medieval period (800-1150 AD). The samples were actually
first discovered by ivory dealers who were using the dense bone to carve knife
handles. Differences in Nitrogen stable isotopes indicate a slight difference
in feeding ecology than the Bering Island population (which makes sense given the geographic separation from
Bering island). They hypothesize that perhaps climatic deterioration combined
with aboriginal hunting by the newly arrived Inuit led to a second, earlier
extirpation of a second historic population of Hydrodamalis.

Pinnipeds are sexually dimorphic - males
are larger than females and are more robust to boot. Most modern pinnipeds are
sexually dimorphic, although it is minimal in most true seals (Phocidae). Extreme
sexual dimorphism characterizes the walrus, all otariids, elephant seals, and
the gray seal. Sexual dimorphism is most extreme in polygynous pinnipeds -
those that breed en masse at rookeries, compete for space, and tend to breed in
harems where competition for mates is strong. How can we study the evolution of
sexual dimorphism? One recent study used only data from modern species only and
promoted an ultimately flawed hypothesis that the earliest pinnipeds were not
only minimally sexually dimorphic but also primitively ice-breeders. This could
have been easily checked using a couple brief readings of the pinniped
paleontological record - none of the earliest known pinnipeds, enaliarctines,
are known from polar regions, and enaliarctines like Pteronarctos goedertae are
already thought to have been sexually dimorphic. The distribution of sexual
dimorphism amongst all three families does suggest that it is primitive for the
group - and many extinct pinnipeds on the stem of extant families (Thalassoleon,
Otariidae; Neotherium, Imagotaria, Dusignathus, Valenictus,
Odobenidae; not sure about extinct phocids) also demonstrate widespread sexual
dimorphism in the past. As with all evolutionary questions, if a question can
be answered with fossils - our only source of objective information about the
history of biological structures - it should. It's actually quite easy, since
sexual dimorphism is reflected in the skeleton - and this new paper by Thomas
Cullen and others does just that. They use a morphometric dataset including a
few extant phocids, extant otariids, the walrus, the extinct pinniped Desmatophoca
oregonensis, and the enaliarctine Enaliarctos emlongi. Two skulls
of E. emlongi are known - the well-preserved male holotype, and a
smaller, crushed skull they reasonably interpret as a female. Their
morphometric analysis (Procrustes principal components analysis) quantifies the
descriptive statements about pinniped dimorphism, and makes several new
morphological observations on dimorphic features; females have been known to
have smaller nuchal and sagittal crests for a long time, but Cullen et al. also
found that they have proportionally narrower palates and rostra, and have a
less robustly constructed squamosal (specifically the mastoid region and the
paroccipital process of the exoccipital). Using this data they executed an
ancestral character state reconstruction, which reconstructs (accurately in my
opinion) sexual dimorphism as being the primitive condition for all pinnipeds.
However, there are a couple issues with this study. First, it is not the first
study to propose that enaliarctines were sexually dimorphic: Annalisa Berta
published evidence for this in Pteronarctos goedertae back in 1994,
though not acknowledged by Cullen et al. Secondly, Cullen et al. only compared
the supposed female skull with Enaliarctos emlongi, E. barnesi,
and E. tedfordi - but not with Pteronarctos (which occurs rather
low in the Newport sequence) and Enaliarctos mitchelli, which is a tiny
narrow-snouted taxon also reported from the Nye Mudstone by Berta in 1991. This
specimen could easily represent another E. mitchelli given its tiny
size, so it is problematic that no comparison was made.

It’s (mostly…) been a good year for marine
vertebrate taphonomy (there are several new marine reptile and whale fall
papers from 2014 that don’t really fit on this list). A few papers over the
last few years have detailed some taphonomic advances on baleen whale skeletons
from the Pliocene of Italy. A new paper by Silvia Danise and Stefano Dominici
takes a quantitative approach towards taphonomic aspects of 25 mysticete
skeletons from shallow marine deposits, mostly from the Pliocene (but one Pleistocene
and one Miocene specimen as well). As discussed above (see Boessenecker et al.
2014), it’s difficult to study modern taphonomic processes on the continental
shelf – currents and rapidly shifting sediment make burial or transport of
carcasses and skeletal elements a certainty, aside from being shallow enough to
permit carcass refloating. Do differences in sediment, fauna, and depositional
processes translate into differences in preservation between the deep sea
(where we have plenty of experimental data, but essentially very few
paleontological examples of true deep sea occurrences) and the continental
shelf (where the sheer majority of marine vertebrate fossils were deposited,
but have virtually no actualistic data)? We should expect differences. Danise
and Dominici report a number of interesting and some previously predicted
patterns – one such predicted example is the lower rate of skeletal
articulation in sandstones as opposed to mudrocks (I reported the same result).
Several headless skeletons were interpreted to be carcasses that had refloated
and disarticulated while floating prior to final sinking, as observed by
aktuopalaontologist Willhelm Schafer in the 1930’s (from a rowboat in the North
Sea). They report a high number of shark teeth associated with skeletons,
including articulated skeletons and (correctly from my perspective) argue that
shark scavenging is ineffective at disarticulating large carcasses (also
confirmed by whale fall experiments) but also making the case that teeth are
only common around skeletons, reflecting scavenging. It’s also possible that
shark teeth are just as common in background sediment, and without looking at
the background sediment, it’s technically not a great argument to make
(although this is me being nitpicky). They also mention bone degradation by
microorganisms or Osedax, but make no mention of the actual traces –
which from the perspective of a reader, does not really eliminate the
possibility that the bones are just poorly preserved or were damaged during
collection or preparation. Several skeletons have associated invertebrates, but
only one has any specialist chemosynthetic (e.g. whale fall) mollusks; the rest
are associated with scavenging or filter feeding mollusks. So, my question is
this: if only one skeleton is demonstrably associated with a fossil whale fall
invertebrate fauna, isn’t only one out of 25 specimens a whale fall?
Regardless, this paper demonstrates that there is a far higher variability in
preservation on the continental shelf – probably reflecting the variation in
lithology, fauna, and depositional processes on the continental shelf. If you
have an interest in marine taphonomy, this is a must-read and one of the most
important studies in this small subdiscipline in years.

How did archaeocete whales hear? We know
that modern odontocetes and mysticetes hear in two different ways: at high and
low frequencies (respectively). High frequency hearing is excellent for
echolocation, as the sounds travel (relatively) slowly and not very far. Low
frequency vocalizations by baleen whales are loud, and travel far and fast –
permitting blue whales to communicate across vast stretches (hundreds of
kilometers) of ocean (although the classic humpback whale song is what most
will think of). Why is hearing frequency so important? One famous baleen whale
individual (yes, a single individual recorded underwater since 1989) of unknown
species – known to the public as the world’s loneliest whale – vocalizes at a
frequency higher than the range of hearing of all other baleen whales,
rendering it incapable of being heard by members of its own species (whichever
one it belongs to). Sadly, this individual travels the globe by itself with
nobody – aside from humans with some microphones – to hear it. But dead whales
sing no songs, so how do you determine the hearing capabilities of extinct
whales? The hearing frequency range can actually be estimated by looking at a
number of features of the inside of the inner ear, including the easiest to
explain example – cochlear coiling. The cochlea is a spiral-shaped organ where
currents in the fluid inside pass over little hairs and are transmitted through
nerves to your brain; the currents in the cochlear fluid come from vibrations
of the stapes, incus, and malleus (the ossicular chain: known informally as the
stirrup, anvil, and hammer, respectively) which vibrate thanks to amplified
sound waves that hit the ear drum. It more or less works in the same general
pattern in cetaceans, with a few differences that need not be explained here –
rather, the take home message is that a more tightly coiled cochlea reflects
low frequency hearing, and a loosely coiled cochlea reflects high frequency
hearing. (Note: all of this happens within the periotic bone which paleocetologists
obsess over so much). Early studies used serially sectioned archaeocete
periotics - serial sectioning is the slow grinding of fossil, with photos or
diagrams recorded incrementally to put together a 3D model and cross section; contemporarily
the results from serial grinding could be called a “bootleg CT scan”. There’s
no excuse for serial grinding anymore, since we have CT technology which (best
of all) actually leaves you with a complete specimen afterwards – after serial
grinding, you’re left with the data you collected and a pile of bone dust.
Serial grinding studies originally suggested that archaeocetes like Zygorhiza were high-frequency adapted.
CT scans of Zygorhiza taken by Eric
and Rachel indicate that hearing was most similar to modern baleen whales – and
that a number of features, including cochlear coiling, are adaptive for low
frequency hearing. This is fascinating and perhaps not surprising from an
ecomorphologic perspective, as the earliest mysticetes were effectively
archaeocetes with broad rostra, whereas contemporaneous Oligocene dolphins like
Cotylocara, Simocetus, and Waipatia
already had most of the adaptations for high frequency hearing and echolocation
present (see Geisler et al., below) – suggesting that whatever the earliest
mysticetes were doing from an ecological perspective, it could not have been
too much different than basilosaurids. Also interesting is the fact that Zygorhiza appears to have been more
sensitive to rapid head movements than baleen whales, suggesting higher agility
amongst basilosaurids.

This study stems from Joe El Adli’s
undergraduate research at San
Diego State University,
which he continued to work on after getting a job at the San Diego Natural
History Museum. The SDNHM had on loan from UCMP a beautiful little
skull of Herpetocetus from the Pliocene San Diego Formation. I was of
course interested in this specimen thanks to my own work on Herpetocetus
bramblei from the Purisima Formation. The San Diego Fm.
sample of Herpetocetus is much larger, and the fossils represent a
somewhat different species than that from the Purisima; the San Diego Formation
is also slightly younger. Herpetocetus morrowi is a small
mysticete, perhaps only 4 meters in total length when alive, with a rather
peculiar jaw joint that would have restricted opening of the mouth to about
15-25 degrees or so – quite the opposite from balaenopterid rorquals, where the
mouth can open to over 90 degrees. Herpetocetus has an archaic braincase
with highly “telescoped” rostral elements, which others have tied to lunge
feeding. However, lunge feeding does not appear to have been possible, owing to
the restricted gape. Instead, this critter has a number of adaptations for
longitudinal twisting of the jaw, in a fashion similar to a gray whale. We make
the case that, based upon anatomical evidence, Herpetocetus was also a
suction/benthic feeder – all other cetaceans with adaptations that limit
oral gape are suction feeders, and longitudinal rotation could be used to
laterally scoop up prey laden sediment into the mouth. This interpretation
dovetails quite nicely with a prior study of mine that reported a middle
Pleistocene extinction of Herpetocetus as well as a study by Nick
Pyenson and Dave Lindberg which identified that glacioeustatic fluctuations in
sea level would have forced gray whales (Eschrichtius) to maintain
alternative feeding strategies (as low sea levels during glacial maxima would
have decreased the amount of available shelf space for benthic prey). Because Herpetocetus
was incapable of lunge feeding and appears to have been overspecialized for
benthic feeding, we hypothesize that increasingly more extreme middle
Pleistocene glacioeustatic sea level changes drove this dwarf baleen whale to
extinction.

Esperante, R., Brand, L.R., Chadwick, A.V.,
and Poma, O. 2014. Taphonomy and paleoenvironmental conditions of deposition of
fossil whales in the diatomaceous sediments of the Miocene/Pliocene Pisco
Formation, southern Peru- a new fossil lagerstatte. Palaeogeography, Palaeoclimatology,Palaeoecology 417:337-370.
Link here
This new paper is the most detailed in a
series of papers by a group of “creation geologists” from Loma Linda University
about the taphonomy of fossil whales in the Pisco Formation of Peru. They
present a rather large body of data collected from field studies in Peru.
Data they report is similar in scope to that reported for marine vertebrates
from the Purisima Formation (Boessenecker et al., 2014, above) with the
exception of having more detailed sedimentologic descriptions. Fossil whales
from the Pisco Formation are frequently articulated and preserved in diatomite,
and occasionally preserved with baleen – which is not surprising, preserved
baleen from these localities has been known since the 1980’s. Notably and
unfortunately, these authors chose not to excavate and collect the fossil
cetaceans: they partially excavated them, and then reburied them. In a way this
makes the work untestable, with the exception that you *could* go out to the
Peruvian desert and dig up another batch of skeletons (on the contrary, every single specimen from Boessenecker et al. currently resides in museum collections so that my results are readily auditable). They conclude based upon
taphonomic (rather than sedimentologic) data that the whales must have been
buried rapidly, and (rather strangely in my opinion) state that Osedax
bioerosion would likely destroy the skeleton if exposed longer than a few
weeks; they cite several modern studies to support this assertion, despite none
reporting complete Osedax degredation of a skeleton in under one year
(and that is only one example; many skeletons at whale falls have persisted for
decades). They do acknowledge that the carcasses could have sunk into soupy
sediment (the precursor to diatomite is called diatomaceous ooze for a reason),
but strangely do not modify their carcass burial rate estimates accordingly.
What I must note is that outside the peer-reviewed scientific literature, these
authors have published creationist articles that extrapolate these problematic
burial rates to the entire Pisco basin, claiming that the basin would have been
filled in a number of millennia rather than over a 15+ million year period as
demonstrated by real dating methods. They then use this to attack absolute
dating methods and claim that radiometric dating does not work. This
information is not exactly hidden deeply on the internet, and one need not
spend much time on google to find out that none of these authors really
believes what they publish in scientific journals. In this context, all sorts
of holes start to appear. For example: if one adopts a disingenuous approach,
why be forthcoming with all of your data? Perhaps the rate of articulation is
exaggerated by the authors only excavating what upon discovery were clearly
articulated skeletons. Where are all the dolphins and pinnipeds? And marine
birds like penguins? The authors hardly make any mention of non-baleen whales –
indicating a bias towards large skeletons. Not all fossil vertebrates in the
Pisco are articulated skeletons – what about isolated elements? Reports of such
occurrences abound. What about fossil material in existing museum collections?
What about bonebeds and layers clearly indicating massive gaps in depositional
history? There is no mention, anywhere, of time-rich bonebeds and preservation
therein. What may look to the uninitiated as a piece of solid science has a
number of problems, and because this publication is being used to push a Young
Earth Creationist paradigm outside the published literature, I cannot
inherently trust its content.

Marine mammals have previously been
reported from upper Pleistocene sediments from upstate New York, Vermont, and Quebec. Anyone who
has visited the famous 18th century star-fort Fort Ticonderoga in
upstate New York would be rather quizzical if you told them that during the end
of the last glaciation, an inland sea occupied that site (and not, for example,
a glacier or a forest). After the last glacial maximum, melt water from the
Laurentide ice sheet sat in an isostatic depression (one formed by compaction
of the earth’s crust due to massive ice loading) and pooled up – forming the Champlain Sea, which was
substantially larger than modern Lake
Champlain and included what is now Lake Ontario.
Marine mammals from Champlain Sea deposits include species now commonly
associated with far colder climates of the Arctic, such as belugas (Delphinapterus
leucas), some of the only known fossils of narwhal (Monodon monoceros),
several cetaceans dubiously identified from vertebrae and ribs (e.g. Phocoena,
Balaenoptera), walrus (Odobenus rosmarus), harp seal (Pagophilus
groenlandica), bearded seal (Erignathus barbatus) as well as
temperate harbor seals (Phoca vitulina). This paper reports a ringed
seal (Phoca hispida) from the Champlain Sea, another
arctic phocid. Other ringed seals have been reported before, so this is not a
surprise, but this new specimen has yielded radiocarbon dates that indicate
marine mammals colonized the Champlain Sea almost immediately after it formed.

Oligocene cetaceans are generally rare
worldwide, but bridge the gap between modern cetaceans and Eocene archaeocetes.
While the archaeocete-mysticete transition is fairly well known, perilously few
early diverging odontocetes have been described. Oligocene odontocetes are not
special by virtue of being Oligocene in age; Waipatia and many other Oligocene
dolphins are members of the Platanistoidea, one of the earliest diverging
dolphin groups with extant members. Other Oligocene dolphins have proven
problematic; Xenorophus and Agorophius are known from the
Oligocene of South Carolina, but the nearly complete skull of Agorophius
has been missing for a century, and Xenorophus is known only from a
partial rostrum. A new extinct dolphin from this same locality - and closely
related to the poorly known Xenorophus - is described in this new paper
as Cotylocara macei. Xenorophids are quite poorly known in the published
literature, but a number of unpublished specimens has meant that many
paleocetologists are familiar with their morphology, despite not being
published; fortunately we no longer have to rely upon unpublished
"conventional wisdom". This new odontocete is spectacularly
preserved, and is held in the collections of the College of Charleston Natural History Museum. Cotylocara has an attenuate and downturned rostrum with at
least 12 double and single rooted teeth (meaning that it is slightly polydont),
large fossae on either side of the rostrum, a blowhole positioned slightly
anterior to the orbits, and an asymmetrical facial region of the skull. It also
has a beautifully preserved but very strange periotic, and is one of the only
odontocetes I've ever seen with an archaeocete-like superior ridge. The
asymmetry of the skull, and phylogenetic position of Cotylocara as one
of the earliest diverging odontocetes, indicates that echolocation in
odontocetes evolved relatively rapidly at or around the Eocene-Oligocene
boundary. Curiously, Cotylocara is also one of the only known
odontocetes to have had a longitudinally twisted rostrum such as recently
reported for basilosaurid archaeocetes by Julia Fahlke and others.

Late Eocene assemblages of marine mammals
are common and geographically widespread; Late Eocene archaeocetes have
near-worldwide distribution. However, early and middle Eocene marine mammals
are rare outside the Tethyan region (Mediterranean Africa, India).
This study reports one of the first marine mammal assemblages from sub-Saharan Africa, collected from
commercially mined phosphatic bonebeds of Togo, a
small country in between Ghana and
Nigeria. Marine mammal remains include fragmentary postcrania of
protosirenid and dugongid sirenians, a new genus and species of protocetid
archaeocete they name Togocetus traversei, and additional isolated teeth
and vertebrae demonstrating the presence of two additional protocetids – one
larger and one smaller than Togocetus. Togocetus is represented
by a large number of isolated bones and teeth, including skull fragments, a
couple of nice mandibular fragments with teeth, and a tympanic bulla. All the
material assigned to Togocetus are of approximately similar size, and
when multiple elements are present, are similar in morphology. This is
admittedly not a very popular manner in which a hypodigm is assembled; ideally,
we would refer specimens that clearly overlapped – but this is a very
tetrapod-specific approach, and generally not followed by shark paleontologists
who must deal with a fossil record which is 99.9% composed of isolated teeth
that must be divided into groups. Regardless, hypodigm assembly such as this
has its merits, and is totally reasonable as it is a testable hypothesis – and
is testable by going out and digging up more fossils.

Beaked whales have all sorts of weird bony
structures of the rostrum, facial region, and mandible, including large paired
fin-like crests (bottlenose whales, Hyperoodon), extraordinarily dense
rostra (Mesoplodon densirostris), and densely ossified mesethmoid
(normally cartilage; Cuvier’s beaked whale, Ziphius cavirostris). It’s
even weirder when you look at fossil ziphiids – some have little bony horns on
the maxillae (e.g. ), a large median ridge or fin of dense bone on the rostrum
(Tusciziphius, Caviziphius), an enormous spherical median nodule
of bone on the rostrum (Globicetus), and longitudinally swollen premaxillae
(Ziphirostrum, Aporotus). Histological study of modern and fossil
ziphiids indicate that many of these different facial ossifications arise via
different pathways, indicating developmental diversity as well as structural
diversity. No single good argument has been made to explain these structures;
in modern ziphiids they are of course sexually dimorphic – so explanations relying
upon dense bone for buoyancy and acoustics (e.g. facilitating hearing) can be
rejected outright, since these structures are absent in females. Rather, they
suggest a behavioral role. Pavel Gol’din put more thought into this, and
suggested that these are auditory “display” structures. These were never
considered for display since they are internal to soft tissue and would not be
visible externally. However, odontocetes can echolocate and experimental data
indicates that dolphins can visualize 3D information from echolocation. Putting
two and two together, Gol’din hypothesizes that these disparate skull
structures would be easily detected from echolocation – and not only species,
but sex would be readily apparent from acoustic visualization (as ziphiid skulls
have robust sexual dimorphism). This in my opinion is a totally fascinating
hypothesis, and one that makes surprisingly good sense.

Archaeocete whales are most famous for
retaining functional hindlimbs, and many early archaeocetes like Pakicetus
and Ambulocetus could walk or run on land – and their appendicular
skeletons looked not too dissimilar from other artiodactyls. Basilosaurids have
reduced hindlimbs and a pelvic girdle decoupled from the vertebral column, and
are thought to have facilitated copulation similar to the vestigial hindlimbs
of boa constrictors. Aside from the singular fact that modern cetaceans retain
a vestigial pelvis (the only reported function of which is to anchor muscles
that move the penis; not sure about function in females), most paleocetologists
who work on Neoceti are unashamedly cranial in focus, something I began to
lament when I started describing postcrania of NZ eomysticetids. The focus on
craniomandibular material has led many neocete workers ignore or give poor
descriptions of postcrania of fossil neocetes. So, what happened in between
having an external hindlimb in the Eocene and a tiny surfboard-shaped pair of
pelvic bones in modern cetaceans? This new paper (also by Pavel Gol’din)
describes several well-preserved innominata (pelves) and partial femora and a
tibia of mid-late Miocene cetotheriid mysticetes from Paratethys (southeastern Ukraine).
These have quite a bit more morphology than the simplified “pelvic surfboard”
of modern cetaceans and permit identification (and clarification) of homologous
features on the cetacean pelvis. These cetotheriid pelves are three-pronged,
and lack a socket for the femur or an obturator foramen (large hole between the
ischium and pubis), and have a posteroventrally descending pubis. Basilosaurid
pelves have an obturator foramen and a socket for the femur (acetabulum), but
the orientation of the innominate is unknown and different interpretations have
been published by Kellogg in the 1930’s and Phil Gingerich in the 1990’s.
Pavel’s study suggests that Kellogg’s early reconstruction – with the hip joint
at the posterior, rather than anterodorsal end of the pelvis – is likely
correct. This also suggests that the left and right innominates may not have
been medially connected at a pelvic symphysis as hypothesized by Gingerich, but
may have been floating in soft tissue as in all modern cetaceans (which is
another point of debate in current archaeocete literature; see recent
publications by Hans Thewissen for more).

This paper is a followup to Gol'din and
Zvonok (2013) who named a new genus and species of basilosaurid archaeocete
whale from the Eocene of Ukraine, Basilotritus uheni. The new genus also
included B. wardii, a species formerly attributed to the protocetid Eocetus.
Basilotritus has distinctive pachyosteosclerotic vertebrae with abundant
external foramina. This new specimen was collected as an associated skeleton
from the upper part of a glauconitic hiatal deposit rich in shark and fish
remains, and includes mandible fragments, numerous teeth, vertebrae, ribs, a
well preserved sternal skeleton, and fragments of the scapula and a single
phalanx. The teeth are similar to other basilosaurids but differ in having
secondary denticles on the accessory denticles (e.g. the denticles on the
postcanine teeth are almost serrated) - a unique feature amongst archaeocetes -
but primitively retain inflated tooth roots like protocetids. As in other
archaeocetes, the sternum is multielement - unlike the single-element sternum
of mysticetes, for example. In general, the dental and skeletal anatomy of this
specimen confirms the transitional morphology of Basilotritus with a mix
of protocetid and derived basilosaurid features and cladistic analysis
demonstrates its position as an early diverging basilosaurid. However, the
cladistic analysis uniquely shows a Neoceti + Ocucajea (another archaic
basilosaurid, from Peru) clade as the earliest diverging lineage within a
paraphyletic basilosauridae, which is surprising given that Dorudon is
typically identified as the sister taxon of Neoceti (albeit, perhaps
erroneously). Better preserved fossils of basilosaurids, and analyses including
a wider range of archaeocetes and neocetes (e.g. not focusing on one with only
a couple taxa from the other) are really needed to further investigate the
archaeocete-neocete transition. Lastly, the teeth show strongly developed wear
facets similar to the protocetid Babiacetus, and perhaps indicative of
predation on sharks as reported for modern killer whales. Gol'din et al.
further mention the abundance of shark teeth in the same horizon as lending support
to this, although since both occur within greensand this is likely
depositionally controlled and simply taphonomic coincidence rather than
anything paleoecologically meaningful.

There has been an explosion of new ideas
and species of “cetotheres” over the past decade, and it seems to be
accelerating. This started with a couple of papers in the mid-2000’s by
Virginie Bouetel who for the first time cladistically demonstrated that a
subgroup of cetotheres (as formerly defined, a totally paraphyletic wastebasket
group) actually do share a number of features that unite them into a clade now
formally (and widely) recognized as the Cetotheriidae sensu stricto (or,
these days, just Cetotheriidae). One fascinating development is the reinvigoration
of study of paratethyan cetotheres. Paratethys was an enormous foreland basin
that was occupied by a continuous inland sea that stretched from Austria
to the Aral Sea in Kazakhstan; the Black Sea, Caspian Sea, and Aral Sea are Holocene remnants of a formerly much larger basin, and
fossiliferous deposits are now uplifted and exposed around their margins and in
between. Brandtocetus is known from a couple of partial skulls collected
from upper Miocene marine rocks on the Crimean peninsula (collected prior to
the Ukrainian civil war, of course). The skull of Brandtocetus is similar in
many regards to Cetotherium, and lacks the twisted temporomandibular
joint of Herpetocetus; the braincase is somewhat wider, and has sinuous
nuchal crests. Most importantly, it has well-preserved earbones that are
removed from the skull, unique amongst described paratethyan cetotheres; in
most other specimens they are still embedded in matrix. This new taxon expands
our knowledge of paratethyan cetaceans, and I was quick to code this new genus
for my own thesis cladistics. Note: it’s been another damn good year for Pavel
Gol’din!

True seals (Phocidae) are the most diverse
group of pinnipeds, and have an extensive fossil record in the North Atlantic,
Mediterranean, and Paratethyan region; however, the record is less robust in
the southern hemisphere and does not include many well-preserved skulls, in
contrast to other groups of pinnipeds in the fossil record. One key phocid is
the extinct Pliocene taxon Homiphoca capensis, known from the early Pliocene
Langebaanweg locality of South Africa. Most of the material is isolated, but only one seal appears to be
present. Additionally, Homiphoca has been identified from the early
Pliocene Yorktown Formation of North Carolina, indicating that it had an
antitropical distribution. The skull of Homiphoca is generalized and
quite similar to the Peruvian phocid Piscophoca. Like most southern
hemisphere phocids, it is a monachine; monk, elephant, and all Antarctic
phocids are monachines – while the Antarctic seals (crabeater, leopard, ross,
and weddell) form the tribe Lobodontini. Homiphoca has been generally
considered to be closely related to other extinct southern hemisphere monachines
like Piscophoca, Hadrokirus, and Acrophoca; however, very
few morphology based cladistic analyses of phocids exist. One of the first good
analyses was published by Eli Amson and Christian de Muizon last year for Hadrokirus
– which recovered Homiphoca, Piscophoca, and Hadrokirus
together within a clade that in turn was sister to the Antarctic lobodontines
and Acrophoca. This new study reports on the cranial morphology and
phylogeny of Homiphoca, using a sample of 7 partial and complete skulls,
figured beautifully in the new paper. Each is coded as a separate OTU in the
analysis to see if Homiphoca capensis truly is a single species. Monophyly
of Langebaanweg seals is confirmed, whereas an earlier morphometric study by
Govender suggested the presence of at least two different species. These new
results suggest that any future splitting of the Langebaanweg Homiphoca may
not be so well-founded, given the poor separation of Homiphoca specimens
in the analysis.The new results, unsurprisingly, place Homiphoca within
the lobodontine clade – but surprisingly - as the sister taxon of the Ross seal
– Ommatophoca rossi – possibly the weirdest looking of all
phocids (internally, anyway – their skulls are very alien-looking). Piscophoca
is placed as sister to the Ommatophoca-Homiphoca clade, with the
other lobodontines and Acrophoca falling just outside, or switching
places with Piscophoca depending upon the analysis. The Pliocene
Langebaanweg assemblage is notable for the abundance of Homiphoca and
complete absence of otariids – today, the cape fur seal Arctocephalus pusillus
is the only local pinniped with Antarctic lobodontines occasionally straying
into south African waters – suggesting faunal changes in the Plio-Pleistocene.
Govender hypothesizes that ancestral lobodontines were primarily accustomed to
hauling out on sandy beaches rather than rocky shores (as modern lobodontines
generally only haul out on smooth pack ice rather than rocky shores), and
dispersed to south Africa via island haul outs; Plio-Pleistocene sea level
transgressions resulted in the proliferation of rocky shores, which otariids
were able to colonize more easily.

Three extant (Boto, Susu, Franciscana)
dolphins and one recently extinct river dolphin (Baiji) are known as river
dolphins because they inhabit rivers (the Francisca actually is marine and
inhabits estuaries, but is anatomically similar and thus grouped with river
dolphins anyway). These dolphins apparently all belong to lineages that
independently “adapted” to freshwater environments, although in most cases it’s
unclear what anatomical adaptations are really needed. Some river dolphins have
smaller eyes, reflecting a stronger use of echolocation and lack of emphasis on
vision in dark, sediment-laden water. One unique adaptation – thought to permit
extra maneuverability in rivers in the Amazon basin – is the “double” shoulder
joint of the Amazon river dolphin, Inia geoffrensis. Aside from the standard
humerus-scapula joint, a second joint is present where the proximal humerus
also articulates with the sternum – which is much wider in Inia than
other odontocetes. Gutstein et al. report an isolated iniid humerus from upper
Miocene nonmarine rocks of Argentina,
which they tentatively identify as belonging to an ischyorhynchine iniid, which
has a proximal end similar to modern Inia. This specimen suggests that
the double shoulder joint adaptation has been around for at least 6 million
years or so.

This short paper reports a single vertebra
they identify as the desmostylian Paleoparadoxia from lower Miocene
rocks of Hokkaido. Paleoparadoxia is abundantly known from Miocene rocks of Japan and
California, and late Oligocene desmostylians are known from both areas (and Alaska) but
include earlier taxa such as Cornwallius, Ashoroa, and Behemotops.
Paleoparadoxia has recently been split up into three genera – Archaeoparadoxia
from the earliest Miocene of Mendocino County, California, and Neoparadoxia
from the late Miocene of Orange County, California; all California
specimens were assigned to these two new genera, with the genus restricted to Paleoparadoxia
tabatai. In this case, the use of the genus Paleoparadoxia in this paper is
now equivalent of Paleoparadoxiinae of Barnes (2013) – who split the genus up.
Hasegawa et al. do cite Barnes (2013), but because I cannot read Japanese, I’m
not sure what stance they take on the splitting. Regardless, this new specimen
demonstrates continuous inhabitation of the western North Pacific by
paleoparadoxiines through the early Miocene, and is one of the first records
from Japan that is temporally equivalent with Archaeoparadoxia weltoni
from California.

Kazár, E., and Hampe, O. 2014. A new
species of Kentriodon (Mammalia, Odontoceti, Delphinoidea) from the
middle/late Miocene of Groß Pampau (Schleswig-Holstein, North Germany). Journal of
Vertebrate Paleontology 34:1216-1230.
This paper reports a fragmentary new fossil
named as a new species of dolphin in the genus Kentriodon. Kentriodon is
known from the middle Miocene of the Atlantic coastal plain in the eastern US,
Sharktooth Hill in the western US, and Japan. This new species, Kentriodon
hoepfneri, is known from a fragmentary skull with a well-preserved periotic
and partial bulla, mandible fragment with teeth, and a nearly complete
vertebral column. Normally I’d be inclined not to treat a specimen like this as
name-able, but the periotic is certainly an exception and importantly preserves
some differences with other Kentriodon, notably a small, oval-shaped facet for
the attachment of the bulla. The Kentriodontidae is another example of a
cetacean family where monophyly is assumed rather than demonstrated, as
correctly noted by Kazár and Hampe; they note that some of the features
identified as supporting this family may be symplesiomorphies (rather, features
that are primitive and cannot really be used to diagnose a clade). Interestingly,
the completeness of the vertebral column permits functional comparisons with
modern odontocetes, and in terms of centrum length profile along the column the
proportions are similar to modern beluga (Delphinapterus) and some
beaked whales (Ziphius, Mesoplodon). The neck and thoracic column
are relatively flexible, while the caudal region is rather stiff; this suggests
a skeleton less adapted to rapid, sustained swimming where the vertebrae have
similarly stiff articulations throughout the column. Lastly, isolated fossil
periotics similar to Kentriodon had previously been reported from
eastern Europe, but this new record robustly confirms the presence of Kentriodon
in the eastern Atlantic, suggestive of a worldwide distribution during the Miocene.

This study is in Japanese (which I am
unable to read) but has an English abstract and excellent figures. This paper
reports a late Miocene baleen whale skeleton from central Honshu, Japan.
The skeleton is rather fragmentary and includes a very partial skull that is
perhaps unusually preserved, well-preserved earbones, and much of a postcranial
skeleton although the bones themselves are rather "chewed up".
Because of this incompleteness, the authors are not able to confidently
identify it to the genus level, and tentatively identify it as a
"cetothere" sensu lato (aka stem Thalassotheria, sensu Bisconti et
al. 2013). The periotic and tympanic bulla, in my opinion, share many features
with archaic balaenopterids like "Megaptera" miocaena
and "Balaenoptera" ryani (both from the upper Miocene Monterey
Formation of California).
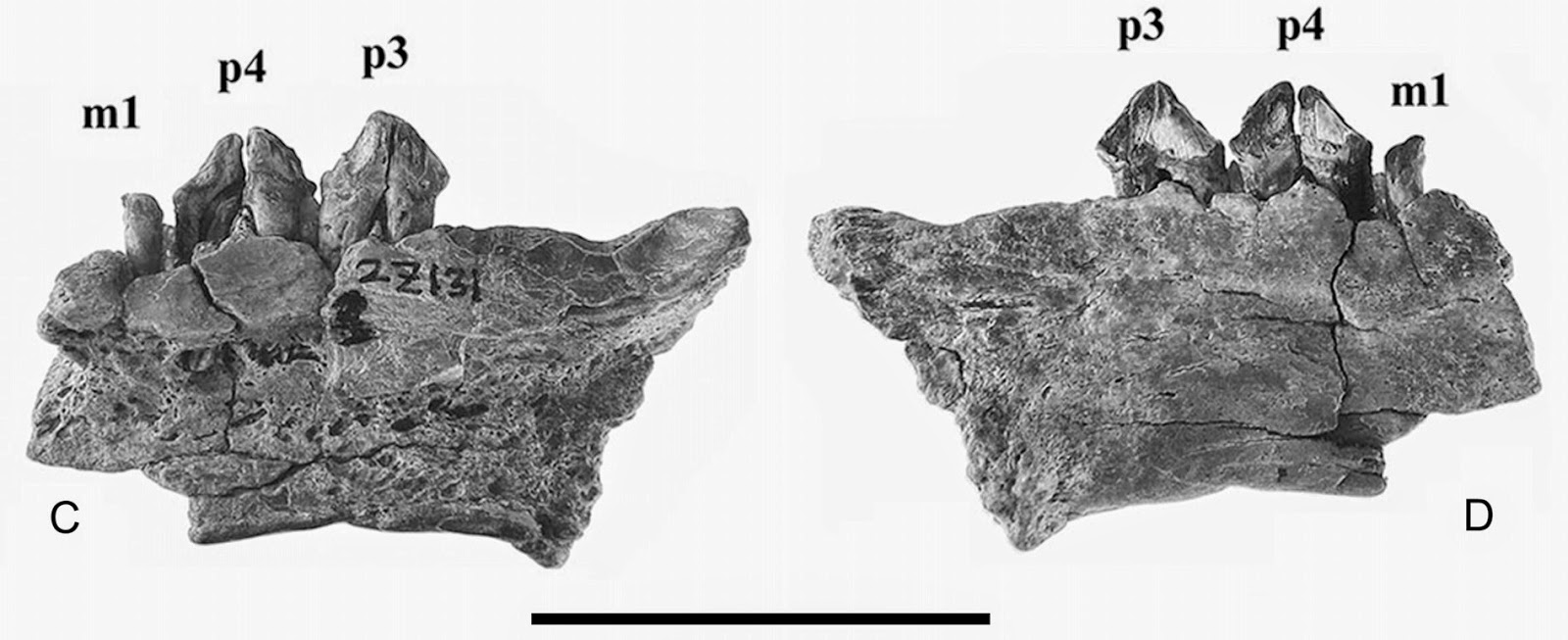
This study reports one of the oldest old
world records of a true seal (family Phocidae), a mandible with a few cheek
teeth which Koretsky and Domning erected the name Afrophoca libyca upon.
The holotype was collected quite recently (2010) by renowned sirenian expert
Daryl Domning from the lower-middle Miocene Marada Formation in the Libyan desert. Based on a
genial tuberosity that extends to the third premolar, they refer the specimen
to the Monachinae. The mandible otherwise has relatively large but simply
ornamented cheek teeth, and reflects a seal similar in size to a modern monk
seal. The locality is 14-19 Ma in age, indeed making this record somewhat older
than most Paratethyan records of seals, and possibly the oldest known record of
any crown pinniped (although Desmatophoca brachycephala from Washington,
USA might actually hold that distinction). Koretsky and Domning point out that
this discovery supports the hypothesis that true seals first arose in the
Oligocene in Paratethys, but did not go so far as to claim that it supports the
mostly passé idea of pinniped diphyly. As an aside, two interesting and
decidedly intentional ‘categorizations’ are worthy of note in the comparisons
section: 1) comparisons with the otter-like putative stem pinniped Puijila
are placed in the “comparisons with Mustelidae” section, and subsequently in
that same section the following statement regarding the dubiously named
“pinniped” Praephoca bellunensis (named by C. Diedrich in 2011): “The
only known specimen, the proximal part of a femur, is so damaged that it is
inadequate for its identification as a pinniped”. Perhaps ironically the same
argument could be applied to similarly preserved partial femora from the
Oligocene South Carolina described by Koretsky and Sanders in 2002.

True seals – also known as earless seals –
have an extensive fossil record in the Atlantic, Paratethys, Mediterranean, and Australasia (and a fragmentary Pacific record limited to the late Pleistocene).
However, unlike the record of otariids (sea lions) and odobenids (walruses),
the phocid fossil record is depauperate with respect to skulls, and as such the
majority of extinct phocid taxonomy is based upon mandibles and “cardinal”
fore- and hindlimb elements of the postcranial skeleton such as the humerus and
femur. An issue with this is that there is sort of a mismatch in the standards
accepted for “otarioid” studies and studies of phocids. For example, although a
couple of walruses have been named based upon postcrania (Pliopedia pacifica and Valenictus imperialensis are the only examples), skulls
and mandibles are generally preferred. This new paper by Irina Koretsky and
colleagues reports three new species of phocids from the upper Miocene Gram
Formation of Denmark based upon fragmentary postcranial elements. The first, Pontophoca
jutlandica, is based upon a partial femur that bears enough similarities to
link it with the middle Miocene paratethyan phocid Pontophoca sarmatica
(Moldavia) – supposedly a monachine. The next, a phocine, is named as Gryphoca
nordica, and is based on a fragment of a proximal humerus – and placed in
the same genus as Gryphoca similis, originally named by Van Beneden from
the late Miocene of the Netherlands and later reported from the early Pliocene Yorktown Formation of
the Lee Creek mine. The last, Platyphoca danica, is also linked with
circum-North Atlantic seal – Platyphoca vulgaris – reported from the early
Pliocene of Belgium and also from the Yorktown Fm at Lee Creek. This
last one is based upon a distal humerus fragment. How distinctive are these
postcranial taxa? Unfortunately not much baseline work exists to evaluate the
likelihood of recognizing the “realness” of these species or confident referral
of additional specimens.

This study is similar in scope to Churchill
et al. (see above), and sought to examine morphological features implicated in
feeding ecology amongst modern and fossil true seals (Phocidae). The study does
present some measurements, but no analysis is attempted and therefore the
findings of the study largely derive from descriptions. Several descriptions of
mandibular morphology are given, including elephant, hooded, gray, spotted,
monk, and crabeater seals, and four extinct seals: Leptophoca, Pliophoca,
Devinophoca, and Miophoca. For some reason dentition is avoided,
and the study focuses only upon the mandible itself. The study identified that
a deep coronoid process and posterior mandible is characteristic of seals that
consume larger prey, while a shallow coronoid process is typical of seals that
eat smaller prey such as fish and krill. Extinct seals like the mid Miocene
phocine Leptophoca lenis (Calvert Formation, Maryland) and the
Pliocene monachine Pliophoca etrusca (Italy) have generalized mandibles
with features of both phocines and monachines – but the rather shallow coronoid
process suggests a diet of mostly small-medium sized prey items. However, since
this study is not quantitative in approach and treats prey size in a
qualitative manner in particular, it’s unclear how meaningful these distinctions
are.

Modern sperm whales are known for their
specialization on eating squid, and the giant sperm whale – Physeter
macrocephalus – is often depicted grappling with giant squid. There is a
hypothesis, though I cannot recall where I read it, that the giant size of Physeter
and the giant squid (Architeuthis) and colossal squid (Mesonychoteuthis)
is driven by an arms race between predator and prey. Unfortunately, we don’t
really have much of a fossil record of soft bodied squid (and none to speak of
for these giants), but the fossil record of sperm whales is robust. Physeter-like
(e.g. subfamily Physeterinae, if you will) teuthivorous sperm whales are known
from the middle Miocene and include critters like Aulophyester and Orycterocetus
– neither of which would have been gigantic, but perhaps the size of a
killer whale. Like modern sperm whales, neither possessed upper teeth, and all
the lower teeth lacked enamel. In contrast, most other Miocene physeteroids
were unlike these derived taxa, and many possessed large teeth with thick
enamel crowns set into robust rostra and mandibles and had skulls with much
larger attachment areas for jaw closing muscles. A ‘killer sperm whale’ habitus
was proposed for late Miocene sperm whales like Zygophyseter and Acrophyseter,
but a macrophagous lifestyle was not completely obvious until the terrifying
monster Livyatan melvillei was discovered in 2010 – a sperm whale with a
3 meter long killer whale like skull (in terms of proportions and robustness)
and teeth the size of 2 liter soda bottles. As a followup to these earlier
studies, this paper reports a new skull of the smaller sperm whale Acrophyseter
that has a series of pathologies on the side of the upper tooth row. These are
a series of bony protrusions (called exostoses) that extend down from the
maxilla and sit on the lateral side of the base of the maxillary teeth. These
are present posteriorly, and are hypothesized (based on similar formation in
better-studied extant mammals like humans) to form as buttresses that support
the teeth during loading. Physics dictates that the teeth closest to the jaw
joint (the posterior teeth) will undergo the highest loading during biting –
and these are exactly the teeth with the exostoses. This study presents
additional data supporting macrophagous behavior in Miocene physeteroids.

This paper describes a new squalodelphinid
dolphin from Peru – from the same unit as the aforementioned Peruvian Notocetus
vanbenedeni (see above – Bianucci et al.), the lower Miocene Chilcatay
Formation. Up until now, the Squalodelphinidae included two key taxa with
complete skulls: Squalodelphis fabianii from the lower Miocene of Italy,
which is known from a complete but otherwise poorly preserved skull with little
surface detail and difficult to see sutures, and the better preserved Notocetus
vanbenedeni from similarly aged rocks in Argentina.
Historically, the monophyly of this group has been assumed rather than
demonstrated, and this study included the first cladistic analysis with
multiple squalodelphinids included – and robustly confirmed – monophyly of this
group (assumptions of monophyly for various “families” remains a bit of a
pervasive problem in paleocetology). Regardless, Huaridelphis is known
by several well-preserved, virtually complete skulls with well-preserved
earbones; this odontocete, like other squalodelphinids, has a relatively
elongate and tapering rostrum, single rooted teeth (aside from some waipatiids,
these cetaceans are some of the earliest odontocetes to evolve single-rooted
dentitions). Subtle differences of the braincase, proportions of various
braincase structures, and the periotics differentiate Huaridelphis from Notocetus
vanbenedeni, as well as having a higher tooth count.

Stenella kabatensis was first described in 1977 by Horikawa from upper Miocene strata
of western Hokkaido, and based on a robust skull that clearly belonged to a delphinid.
Subsequent workers doubted attribution to Stenella, which is a small and
relatively gracile modern delphinid (Spinner dolphins, Stenella longirostris,
are a prime example); Waseda University Ph.D. student Mizuki Murakami set out
to redescribe this problematic fossil as part of his dissertation. Previous
cladistic analysis of delphinoids by Murakami et al. (2013A,B) suggested that S.
kabatensis was much lower on the cladogram; additionally, S. kabatensis
is 8-13 Ma in age, much older than the inferred early Pliocene molecular divergence
date of the Stenella species complex. As it happens, S. kabatensis lacks
a bunch of features found in derived delphinids, and actually shares a few
features of the facial region, mandible, and hyoid apparatus with killer whales
(Orcinus orca) and the problematic fossil delphinid Hemisyntrachelus cortesii
(debate continues as to whether Hemisyntrachelus is distinct from
bottlenose dolphins, and whether all the species contained in Hemisyntrachelus
should just be transferred to Tursiops instead; Murakami et al.'s
results suggest that Hemisyntrachelus is in fact distinctive). Because
of these differences in morphology and concomitant different placement on the
delphinid tree, Murakami et al. erect the new genus Eodelphis, and note
that this dolphin is now the oldest known true oceanic dolphin (Delphinidae).
For years an undescribed skull briefly mentioned in an old paper from the
Monterey Formation of Orange County, California, was accepted as the oldest
known delphinid - but because it was not published, its identification as a
delphinid cannot really be formally audited. Murakami et al. further note that
all Miocene delphinids have been reported from the Pacific (whilst being absent
from richly fossiliferous and densely sampled assemblages from the Atlantic and beyond), strongly
suggesting a Pacific center of origin for the clade. However, molecular results
suggest a separate origin outside the Pacific - discovery of more fossils is
needed to further evaluate the center of origin for Delphinidae. This study
actually became the featured article for the May 2014 issue of JVP, and I was
honored to make the cover illustration (see image below). Thanks again to my
friend and colleague Mizuki for inviting me to contribute!

Despite Eodelphis being a
replacement name for "Stenella" kabatensis, the genus
name Eodelphis has been occupied for nearly a century for the well-known
marsupials Eodelphis cutleri and Eodelphis browni. See, the root
"delphis" also applies to opossums - for example, the Virginia
Opossum is Didelphis. So, Murakami et al. quickly published this
followup note proposing the second new genus name Eodelphinus for the
Japanese fossil dolphin.

Modern toothed whales are notable for
conspicuous skeletal and facial left-right asymmetry, which is associated with
echolocation. Soft tissues in the facial region associated with the nasal
passages have been modified to produce all sorts of sounds, notably clicks
(used in biosonar) and all sorts of whistles. The prevailing hypothesis is that
the right nasal passage and associated musculature has been modified for sound
production while the left nasal passage is less modified, and perhaps preferred
for respiration. As a result, the maxilla and premaxilla are often wider on the
right side than the left, effectively making the midline of the skull shift to
the right; the vertex (top of the skull) is similarly shifted to the left hand
side. Most Oligocene odontocetes have facial regions that are more or less
symmetrical (or, about as symmetrical as terrestrial mammals), but Pliocene and
many middle-late Miocene odontocetes have directional asymmetry in the skull.
Asymmetry and twisting of the rostrum in archaeocetes (and now, the xenorophid
dolphin Cotylocara) is probably not related to echolocation, however.
When did asymmetry arise? Did it arise once, or independently several times?
Larry Barnes hypothesized that facial soft tissue asymmetry is probably a
uniting feature of Odontoceti: all extant odontocetes, with or without cranial
asymmetry, bear asymmetrical soft tissues. Thus, Barnes proposed that
odontocetes with symmetrical crania likely had asymmetrical soft tissues for
echolocation, with cranial asymmetry arising multiple times where soft tissues
became modified enough to influence asymmetry in the skull. This new study by
Mizuki Murakami reports a partial odontocete braincase representing one of the
earliest records of a delphinoid worldwide – a kentriodontid skull from the
lower Miocene of Japan. The specimen clearly has asymmetrical nasals, and the
midline is drawn onto the specimen by the authors; measurements of the distance
of various features from the midline are used to determine the extent of
asymmetry. However, the left and right edges of the skull are missing, and it
is perhaps not possible to objectively conclude where the midline actually
lies. The skull is certifiably asymmetric based upon the nasals – but the
placement of the midline is problematic given the incompleteness of the fossil.
However, the ancestral character state analysis reconstructed the ancestral
delphinoid as having a symmetrical skull – reinforcing the hypothesis of
Barnes.

This new study reports a spectacular new
assemblage of marine mammals in upper Miocene sandstones from the Atacama desert near Caldera in Chile.
These deposits are equivalent in age to some of the younger levels of the Pisco
Formation in Peru, and has even produced marine mammals previously known from Peru such
as the walrus faced dolphin Odobenocetops and the aquatic sloth Thalassocnus.
However, the star of the new paper are not these weirdos but a series of
articulated rorqual skeletons (balaenopterids – e.g. the family that includes
minke, fin, blue, and humpback whales). Articulated skeletons are concentrated
into four horizons. Sedimentologic evidence suggests that this was a sort of supratidal
flat with abundant algae within an embayment. The concentration of these
skeletons, both vertically, and laterally (many of the skeletons are adjacent)
suggests that mass death events at sea resulted in carcasses drifting into this
embayment to find their final resting place on the tidal flats – perhaps killed
off by an algal bloom. I’m very skeptical of claims of mass death assemblages
in the marine vertebrate record since there is such an enormous taphonomic
overprint that usually obliterates any genuine ecological signal in the fossil
record, but in this case the authors have done a fine job that withstands
robust scrutiny.

This paper reports the strangest marine
mammal described this year – the bizarre prognathous porpoise Semirostrum
ceruttii from the Pliocene San Diego and Purisima Formations of
California. Semirostrum is a true porpoise (Phocoenidae) and intermediate
in size between bottlenose dolphins and smaller harbor porpoises. It had a
mostly symmetrical skull with large premaxillary eminences (a phocoenid
feature), and the skull is otherwise unremarkable; all the action is in the
lower jaw (mandible). The mandibles are fused at the symphysis (=chin), and the
symphysis is dorsoventrally expanded into a toothless paddle shape – the jaw is
strongly prognathous, meaning that it extends beyond the tip of the rostrum.
This is somewhat reminiscent of the eurhinodelphinids, which were instead
extremely retrognathous (=overbite), and had a toothless rod-like rostrum that
extended far beyond the tip of the mandible. What could this bizarre structure
have functioned as? The symphyseal region has a number of longitudinal canals,
which could have housed arteries for a keratinous sheathe or other soft tissue
structure, or nerves to permit strong tactile perception (touch).
Interestingly, the teeth have strongly developed wear facets but only on the
labial (lip/outer) side of the teeth, which rather than developing from wear
against other teeth, appears to have been caused by interaction with a foreign
substance. We interpreted this toothwear as being caused by sediment abrasion,
and that Semirostrum used its strange chin to probe, plow, or stir
through sediment looking for soft bodied invertebrates. Notably, several other
benthic feeding marine mammals are known from the Pliocene of California,
including the toothless walrus Valenictus, one other odobenine walrus,
the dwarf whale Herpetocetus morrowi (see above – El Adli et al., 2014),
and an unnamed genus of gray whale (Eschrichtiidae). Lastly, I'll note that the
reason I was attached to the paper was to include several additional specimens
of Semirostrum from the Purisima Formation of northern/central California (Santa Cruz, Half Moon Bay regions) -
material collected by myself and dogged amateur paleontologist Stan Jarocki of Watsonville. One
skull from the Purisima is somewhat older and perhaps represents a second,
older species as it retains obvious facial asymmetry (a primitive trait amongst
phocoenids).

Modern odontocetes have all sorts of
strange holes in their heads - technically known as sinuses. The most well
known sinuses surround the tympanoperiotic, and are thought to acoustically
isolate and insulate the inner ear from bone-conducted sound to permit
directional hearing underwater. Odontocetes - particularly delphinoids - have a
lobe called the pterygoid sinus which extends anteriorly and wraps around and
in front of the nasal passages. Phocoenid porpoises have a lobe that even
invades between the maxilla and frontal bones in the face; these sinuses are
air-filled, causing a huge mismatch in density that is known as acoustic
impedence. It's difficult to study these, since these are cavities within a
skull - in the old days, skulls would have to be sawed apart to see inside.
Nowadays though, CT scanning permits non-destructive 3D imaging of the insides
of the skull. My colleague Rachel Racicot has carved out a niche doing this for
modern and fossil cetaceans. In addition to the sinuses, CT data can also
reconstruct the brain from fossil skulls. This study reports CT findings of the
brain and sinus endocasts of the recently described skimmer porpoise Semirostrum
ceruttii. Semirostrum does not differ in its cranial nerves or cranial
circulatory system from modern porpoises, but bears pterygoid sinus features of
both porpoises and delphinids (attesting to its position as a stem phocoenid),
and differs from most delphinoids and river dolphins in possessing an unusually
robust ossified falx cerebri that penetrates deeply between the cerebral
hemispheres. Racicot and Rowe hypothesize that this may be an adaptation
towards reducing inertia of soft tissues during rapid movements of the head or
perhaps deep diving - whatever the function, it does imply a different sort of
behavior for Semirostrum relative to other delphinoids, as we (see
Racicot et al., above) hypothesized for this bizarre porpoise.

Kentriodontids are a group of early
delphinoids implicated in the ancestry of modern porpoises (Phocoenidae),
oceanic dolphins (Delphinidae) and belugas/narhwals (Monodontidae); their
fossil record extends back to the early Miocene (see Murakami et al., above)
and they more or less have a worldwide distribution (both coasts of North
America, Japan, South America, New Zealand, Europe; see Kazár and Hampe,
above). The most well preserved and well known kentriodontids are all from the
east coast of the United States, principally from the Calvert, Choptank, and
St. Marys formations - small dolphins like Kentriodon pernix, Liolithax
pappus, slightly larger beasts like Lophocetus calvertensis and Hadrodelphis
calvertensis, and the gigantic dolphin Macrokentriodon morani. The
monophyly of this "family" has in the past generally been assumed
rather than demonstrated, although a few studies like Murakami et al. (2014 -
above, on the asymmetric kentriodontid from Hokkaido) have supported
monophyly of a subset. In the 1930's, Kellogg named a bunch of kentriodontids
based upon isolated periotics from the middle Miocene Sharktooth Hill Bonebed
of California (not very good practice) which turned into a bit of a mess once
the first good kentriodontid skulls were being discovered there. Part of this
mess was mopped up by Barnes and Mitchell in their 1984 reevaluation of Kentriodon
obscurus, to which they referred a skull - the species was originally
named Grypolithax obscura, which Barnes and Mitchell thought was
very close to Kentriodon pernix, and a new skull confirmed the
presence of Kentriodon from Sharktooth Hill; the species was recombined
and the skull referred to the species. With that overly long intro out of the
way, this new paper reports one of the first new kentriodontid specimens from
the west coast reported in decades - a well-preserved skull with associated
mandible fragments, teeth, vertebrae, and ribs (sadly, no periotic is
preserved). The new dolphin, named Kentriodon diusinus, is from
the middle Miocene Rosarito Beach Formation near La Mision in northwestern Baja California -
a well known locality notable for having a number of species closely related to
those from Sharktooth Hill. Kentriodon diusinus is relatively
small and has a tapering rostrum with single rooted, conical, polydont teeth,
and relatively large and elongate pterygoid sinuses on the basicranium that
extend very far anteriorly, lending the dolphin the species name diusinus.
Phylogenetic analysis indicates that Kentriodon diusinus is most
closely related to the Sharktooth Hill taxon Kentriodon obscurus;
interestingly, the specimen has previously been identified in abstracts as Kentriodon
sp., cf. K. obscurus. Many more spectacular marine mammal fossils from
the Oligocene and Neogene of Baja California await publication, and I'm pleased
to see this published.

This study (in Japanese) reports taphonomic
details of a whale skeleton collected from the Pisco Formation in Peru. The
fossil was originally collected by the Black Hills Institute, and presumably
purchased by the Gunma Museum of Natural History; I remember seeing the
specimen on the BHI website and hoping it would end up in a museum. The
specimen is spectacularly preserved and represents an archaic balaenopterid
like Protororqualus or “Balaenoptera” cortesi var. portisi.
Associated with the skeleton are 16 shark teeth all belonging to the extinct
“mako” Carcharodon hastalis. Furthermore, the fact that at least two
teeth are from the same tooth position, indicating that at least two sharks
likely fed on the carcass and shed teeth during feeding.

A partial skull and postcranial skeleton of
a small odontocete was found at the Trig Z locality in the Waitaki Valley, a
region well known for its upper Oligocene marine vertebrates. This specimen was
collected in 1954 by T.G. Marples, the son of Otago Zoology professor Brian
Marples (who collected the above mentioned material of Tohoraata
waitakiensis; see Boessenecker and Fordyce 2014A, above), and named Prosqualodon
marplesi in his honor as part of an honor’s thesis by Mel Dickson in 1964.
Historically, Prosqualodon has been a bit of a garbage can; two
correctly referred species exist, including Prosqualodon australis from Argentina
and Prosqualodon davidis from Australia
(the holotype of which has been lost). In the 1930’s, Professor Benham
described another odontocete from Otago – probably from the lower Miocene
Caversham Sandstone not far off from Dunedin itself (the town where our
university is located) – and named it Prosqualodon hamiltoni. This thing
is enormous, and does represent a squalodontid, but is clearly much larger than
Prosqualodon and had a longer rostrum, and is also needing
redescription. Prosqualodon marplesi on the other hand is perhaps more
obviously not a squalodontid, and in 1994 R.E. Fordyce recombined it as Notocetus
marplesi. Years later, it became evident that this was also incorrect, and
as part of Yoshi Tanaka’s Ph.D. thesis he redescribed P. marplesi.
Confirming using cladistic analysis that it did not belong in the genus Notocetus,
he erected a new genus for it – Otekaikea. Cladistic analysis also supported
a sister taxon relationship between Otekaikea and Waipatia, and
thus the new genus represents a waipatiid rather than a squalodelphinid (as it
was reinterpreted by Fordyce, 1994). Otekaikea and Waipatia are
separated only be some rather subtle differences in the braincase and periotic,
which I will abstain from recalling to please non-odontocete interested
readers. Cladistic analysis also indicates that the Squalodelphinidae –
previously assumed to be a clade – is possibly paraphyletic (although see Lambert
et al. 2014, who found squalodelphinid monophyly, but used a smaller matrix).

This is yet another report of a new
cetotheriid from Paratethys. This new mysticete, Zygiocetus nartorum,
was previously referred to as "Cetotherium sp." in an earlier
publication by Tarasenko, and is known from similarly aged upper Miocene marine
rocks from Adygea (Adygea is a small region of the Russian Federation, and lies
to the southeast of Ukraine in the western Caucasus). Most of the features that
actually differentiate it from Cetotherium are relatively minor and
include subtle differences in the nuchal crests on the braincase and
differences in the shape of the periotic. I'm not necessarily certain that
these differences merit separation at the genus level. Unfortunately, the
figures are somewhat spartan and it's not easy to interpret aspects of the
tympanoperiotic morphology. The article oversimplifies patterns of cranial
telescoping amongst mysticetes, claiming to identify a new "type" of
telescoping; in fact, the different reported patterns have been widely
recognized by mysticete workers for nearly a century (to say nothing of the
lack of remarks concerned with telescoping in balaenids, eschrichtiids,
eomysticetids, and toothed mysticetes).

Ontogeny is a confounding problem in
paleontology (see below), but it also can help us understand how certain
morphologies evolve. A classic research focus in evolutionary biology is the
study of heterochrony – changes in ontogenetic timing. Most evolutionary
changes are thought to arise by certain differences in timing. For example,
many features evolve simply by appearing slightly earlier during ontogeny
during later generations – a classic example would be the gradual loss of
abundant body hair in the human lineage. Less hair is common in hominoid
juveniles, and in humans the appearance of hair has been delayed later and
later into ontogeny, and hairless stages of prenatal ontogeny have been delayed
well after birth. Can heterochrony explain some of the morphological diversity
and evolution of baleen whales? As part of his Ph.D., my labmate Tsai conducted
a geometric morphometric analysis (using relative warps, IIRC) of mysticete
ontogeny, focusing on the pygmy right whale Caperea marginata, the sei
whale Balaenoptera borealis, and the humpback whale Megaptera
novaeangliae. The analysis indicated that only minimal changes occur during
the ontogeny of Caperea, with juvenile and adult specimens plotting in
the same region of morphospace. However, adult Balaenoptera and Megaptera
plotted closely, with juveniles of each plotting close together but not
with the adults – indicating not only more extreme ontogenetic changes
(generally reflecting the extreme lengthening of the rostrum in rorquals) in
rorquals relative to Caperea, but similar ontogenetic trajectories in
each rorqual. Tsai and Fordyce thus robustly identify Caperea as
undergoing paedomorphosis – retention of juvenile features into adulthood –
whereas the two rorquals undergo substantially more changes during ontogeny
than the hypothetical ancestral mysticete, and thus undergo peramorphosis.
You and I, and almost all adult humans
(barring rare developmental disorders) do not look like we did when we were
babies, juveniles, or even awkward teenagers. This is mostly true of all
mammals; as mammals mature – some faster than others – a series of skeletal
changes accompany all sorts of other soft tissue anatomical and behavioral
changes. Many mammals are born with relatively short snouts which then
lengthen. In some cases, juveniles of certain species resemble one another
closely enough that species differences are not obvious until maturity (a
common phenomenon in birds). Biologists have the luxury of tracking a single
individual or watching juveniles interact with adults or actually keeping a
captive individual to watch it grow and see how it changes; indeed, ontogenetic
changes are so easily interpreted that most birding guides will show pictures
of juveniles alongside adults for quick visual identification. But
paleontologists are not so lucky; most fossils don’t have ancient DNA to sample and confirm
relationships, and certainly all fossils are dead, forever. How can we tell a
juvenile from an adult? Occasionally size is used – but some paleontologists
assume that juveniles will look identical to the adult, and assume a smaller
specimen with a few slightly different features from a larger specimen might
instead represent a new dwarf species that is closely related. In dinosaur
paleontology, a number of paleontologists have gotten into trouble for naming
new species that other researchers have claimed actually represent ontogenetic
synonyms. How do we assess ontogeny in fossils? We can use paleohistology;
others suggest that you can use cladistics, and that if a potential new species
plots separately, then it truly is separate – whether juvenile or not. Some
work on hadrosaurid dinosaurs has indicated that when known juveniles are coded
they still plot out down the tree away from the adults, with other juveniles.
In mammals it’s a bit easier because we have teeth, and only one adult set –
but baleen whales do not, posing a problem not unlike that facing dinosaur
workers. With this in mind, my labmate Tsai took juvenile and adult specimens
of modern species of baleen whales and coded them separately and ran it in two different
previously published datasets; he used a sei whale (Balaenoptera borealis)
and a pygmy right whale (Caperea marginata). Caperea – which is
paedomorphic with similar adult and juveniles – plotted together, indicating
ontogeny is not much of a confounding factor. However, the juvenile Balaenoptera
plotted well outside Balaenopteridae, and outside Crown Mysticeti
(Balaenomorpha) in one analysis and with Balaenidae (right whales) in the
other. Why? Balaenopterids undergo a number of cranial transformations during
ontogeny, and the morphological gulf between juvenile and adult is far, far
greater than for the pygmy right whale. Few fossil mysticetes for which fossil
ontogenies are known are like Caperea, and thus the potential for being
misled by cladistics and misnaming of juveniles as new species in the fossil
record is great. Possible remedies include not basing new names on juveniles,
unless you can demonstrate that some of the diagnostic features do not change
during ontogeny.

Marine mammals from the Pleistocene are
rare - for most of the Pleistocene, sea level was lower - meaning that most
Pleistocene marine vertebrate bearing localities are below sea level.
Occasionally we get areas where dredging yields plenty of marine mammal fossils
- and hot spots such as this include the North Sea, offshore Portugal and
Spain, offshore South Africa, and the Penghu channel immediately west of
Taiwan. Taiwan has a limited but crucial marine mammal record - few other
localities are as close to the equator, and it's the lowest latitude marine
mammal locality in the western North Pacific; the next richly fossiliferous
localities to the south are in Australia
and New Zealand. The majority of fossils from the Penghu channel are terrestrial
mammals, but various odontocetes have been dredged - and this report describes
two immature braincases of gray whales, Eschrichtius sp. Gray whales are
perhaps most famous for being commonly visible in the spring along the western
US where they are easily seen migrating along the coast (on a drive north from
Los Angeles, my wife and I counted 40+ gray whale spouts (aka blow) in under
two hours of driving through Big Sur). However, few outside cetology are
readily aware that 1) gray whales were declared extinct and rediscovered, 2)
gray whales used to exist in the North Atlantic but have been extinct in Europe
since about 500 AD and the east coast of North America since about 1700, and 3)
still exist in the western Pacific (albeit with a much smaller population).
Gray whales undertake marathon migrations from their calving ground in Baja California to
the Bering Sea in the eastern Pacific, and from the waters off northern China and
Japan to the sea of Okhotsk in the western Pacific. The location of the western population's
calving ground is unknown, but thought to be somewhere around the coast of China.
The presence of two juvenile specimens of Pleistocene age suggests that perhaps
the breeding ground was at least as far south as southern Taiwan,
and possibly even further south.

The postcranial skeleton of archaeocetes is
famous for demonstrating a beautiful transition from the largely terrestrial
mammalian bodyplan to the highly modified skeleton of “modern” cetaceans; key
transitions include the loss of weight bearing ability in the fore- and
hind-limbs, “locking” of the elbow, modification of carpals into blocky
elements and planar interphalangeal and metacarpal joints for stiffening of the
carpus, loss of hooflike ungual phalanges, loss of fusion of the sacral
vertebrae and decrease in number of functional sacral vertebrae, decoupling of
pelvis from vertebral column, reduction of the hindlimb, increase in number of
postcervical vertebrae, and modification of the terminal caudal series for the
caudal fluke and peduncle. Excellent skeletons of some protocetids exist (Rodhocetus,
Artiocetus, Maiacetus) but detailed descriptions have yet to be
published; good descriptions exist for the holotype skeleton of Ambulocetus
and postcrania of the pakicetids Pakicetus and Ichthyolestes, but
these are all isolated elements from a bonebed and some uncertainty exists as
to their allocation. Regardless, protocetids represent the crucial stage
between mostly terrestrial and fully marine cetaceans. A new skeleton of the
protocetid Natchitochia jonesi from the middle Eocene permits a more
detailed assessment of the pelvic evolution in protocetids. Described earlier
from an associated set of vertebrae, this new specimen is a match but also
possesses part of a hindlimb including a well preserved innominate and a femur.
The innominate of Natchitochia dwarfs that of Georgiacetus and
most other protocetids, and the vertebrae indicate it had two sacral vertebrae
(as opposed to four, the primitive condition amongst artiodactyls). In
comparison, Georgiacetus is thought to have zero functional sacral
vertebrae (like basilosaurids). In 2008 Uhen referred some postcrania to Georgiacetus
that suggested that it retained robust hindlimbs but had a pelvis decoupled
from the vertebral column; this new specimen of Natchitochia has a
robust femur and innominate, but unfused sacral vertebrae with a minimal
connection to the pelvis. This new specimen thus supports the idea that
protocetids utilized dorsoventral undulations of the hindlimbs rather than the
tail for aquatic propulsion – however, see Bajpai and Thewissen above
(although, it’s important to note that the protocetid postcrania reported by
Bajpai and Thewissen lack hindlimb elements, so it is not objectively possible
to compare the relative importance of the tail versus the hindlimb in
locomotion – unlike Natchitochia).

Protocetids are a group of archaeocetes
that were both arguably the first oceangoing cetaceans, and also the last group
capable of hauling out on land – perhaps equivalent to the current stage of
evolution of seals and sea lions. Protocetids are well known from the
southeastern USA, and include important taxa like Georgiacetus and Carolinacetus,
which have informed us greatly on the locomotion and ear anatomy of
protocetids. This new paper by Mark Uhen reports several new occurrences of
protocetid teeth from New Jersey and South Carolina. Specimens from New Jersey were actually collected from a Miocene
unit alongside many obvious examples of Miocene terrestrial mammals, leading
Uhen to interpret these as reworked teeth, since protocetids are not known
anywhere else to be younger than middle Eocene. These specimens are not
anatomically revealing, but importantly the New Jersey
specimens reflect the farthest north that protocetids have been discovered.
Uhen suggested that protocetids were confined to the Tethys and north Atlantic owing to their absence
in other well-sampled Eocene rocks from other regions. However, further
sampling in places like Peru, New Zealand, and Seymour Island (Antarctica) is probably necessary. Several protocetids were reported from
fragmentary remains by Uhen et al. in 2011 but are apparently now regarded as
basilosaurids (they are similar to “Eocetus”, one species of which – Eocetus
wardii – was transferred to the new archaic basilosaurid genus Basilotritus
last year).

This solid new study by Jorge Vélez-Juarbe
and Daryl Domning reports a new Oligocene sirenian from the southeastern United States, in the ninth paper in the long-standing Atlantic and Caribbean fossil sirenian
series. The genus Metaxytherium is a speciose grade of sea cows formerly
known only from the Miocene and earliest Pliocene. Fossils of Metaxytherium are
known from all over the world, including the western and eastern North Atlantic, the eastern
North Pacific, the Mediterranean, Indian Ocean, and western South Atlantic. The Metaxytherium “lineage” is paraphyletic, and the
Hydrodamalinae (Dusisiren + Hydrodamalis – the giant sea cows) is
nested within (and of course, the genus Dusisiren is just as
paraphyletic). The new species, Metaxytherium albifontanum, extends the
fossil record of the genus back into the Oligocene. This new species is rather
small, only 2-3 meters in length, and is known from three well-preserved
partial skeletons from the Parachucla Formation of Florida and the Chandler
Bridge Formation of South Carolina (the latter is the same unit which produced
the holotype specimens of notable fossil cetaceans such as Agorophius, Cotylocara,
Eomysticetus, Micromysticetus, and Xenorophus).
Interestingly, this new species of Metaxytherium was apparently
sympatric with other sirenians such as Crenatosiren olseni and Dioplotherium
manigaulti, and because it had smaller tusks and a less deflected rostrum, M.
albifontanum likely relied less upon uprooting rhizomes from the sediment
than these other sirenians. Lastly, this new species suggests that a west
Atlantic origin for the Metaxytherium lineage is likely, rather than a
Tethyan origin.

And this study marks the tenth contribution
in the long standing Atlantic and Caribbean Sirenia series, and Jorge’s second installment as
first author. This study reports a larger sirenian, the holotype of which was
collected by none other than the first author himself in 2006 and 2007 while
doing fieldwork in Puerto Rico. I remember seeing occasional photos of the specimen as Jorge was
preparing it; it is more than admirable for a paleontologist to have the
necessary skills to excavate, prepare, and publish their own fossils (in fact,
that is the very definition of a paleontologist, and unfortunately many
researchers do only the latter). The type specimen of Priscosiren atlantica
is from the San Sebastián Formation and is early Oligocene in age; several
additional specimens were referred from the same unit, and critically, also
from the Ashley Formation of South Carolina. Priscosiren is a
halitheriine dugongid, and other cladistic analyses by Jorge and Daryl have
confirmed its position as a basal dugongid. Priscosiren was likely
sympatric with other Oligocene sirenians such as Caribosiren and Crenatosiren
(being somewhat older than the late Oligocene sirenian assemblage from the
Chandler Bridge Fm. – e.g. Metaxytherium, Crenatosiren, and Dioplotherium).
Priscosiren had a poorly deflected rostrum with small tusks (not
dissimilar from Metaxytherium albifontanum), and as in all known
sirenian assemblages there are subtle differences in feeding morphology between
each sirenian present – but without a clear, consistent pattern through earth history,
suggesting these are “dispersal assemblages” or represent “chance structuring”.
Another solid piece on sirenians, with beautiful color images to boot!

Morocco is rather famous for a series of spectacular marine vertebrate
fossil assemblages spanning the late Cretaceous through Eocene – many fossils
of mosasaurs, and Paleocene-Eocene shark teeth are easily found for sale. In
fact, K-Pg shark teeth from Morocco
can be found in virtually every rock shop in the United States, and in every country I’ve ever been in, including the UK and New Zealand, I have seen them for sale. Normally when abundant sharks are
found, other marine vertebrates are easily recovered; while the Moroccan record
of mosasaurs is great for the Cretaceous, but unfortunately not much in the way
of marine mammals have been found in Eocene rocks. My colleague Rachel Racicot
(Postdoctoral fellow at Howard U.) went to these coastal localities with
archaeocete specialist Mark Uhen (George Mason U.) and didn’t turn up much; the
localities are pretty heavily picked over and quarried by commercial
collectors. Still, some tantalizing bits have been found, and this new paper
reports some of the first Eocene marine mammal assemblages from Morocco.
Cetaceans include a host of basilosaurid archaeocetes, including cf. Saghacetus
sp. (rostrum fragment with teeth, vertebrae), cf. Stromerius sp.
(vertebrae, humerus), cf. Dorudon atrox (vertebrae), cf. Dorudon
sp. (vertebrae, phalanx), and Basilosaurus isis (partial
articulated vertebral column); the sirenian cf. Eosiren sp. (ribs) is
also present. Fossil vertebrate material is concentrated into two bonebeds
separated by a few meters of rhythmic estuarine deposits. The two bonebeds are
for some odd reason correlated with two sea level low stands – which is
curious, since marine bonebeds are typically formed during transgressions (e.g.
see Boessenecker et al., above), and both bonebeds mark deposits deeper than
the intervening estuarine deposits.
1 comment:
Aloha Bobby. It is always a pleasure to read your blog. And this is hopefully an interesting link for the ontogeny difficulty.
Good luck for your PhD!
Indira
http://onlinelibrary.wiley.com/doi/10.1002/jmor.20288/abstract
Post a Comment