
A return to my previously abandoned series on lesser known controversial topics and disagreements in Cenozoic marine vertebrate paleontology - I avoid topics that I am personally opinionated about but am the only critic, as I don't want to use this blog as an alternative to publishing my own opinions. Instead, this series aims to summarize published issues between working groups. This one covers a fascinating new study and two comments/replies about a purported extinction in pelagic sharks discovered in observations of shark denticles (scales) in deep sea core samples.
Introduction
Last year (2021) Elizabeth Sibert (Harvard, Massachusetts), and coauthor Leah Rubin (College of the Atlantic, Bar Harbor, Maine) published a surprisingly provocative paper on assemblages of shark denticles from the deep sea floor. Sibert has published some pretty incredible papers over the past few years applying methods typically employed by micropaleontologists studying fossil plankton – typically critters like foraminifera – to shark and ray denticles from deep sea core samples. Deep sea cores are a critical source of biostratigraphic data for the marine record, especially for the Cenozoic – in my own research I am frequently double checking microfossil chronologies based on deep sea core samples. Core samples, for the uninitiated, are cylinders of sediment and rock drilled by a special circular drill bit. These provide a record of plankton faunal change but also can be studied from a paleomagnetic perspective and also yield a wealth of isotopic data that can be used to trace the history of climate change (e.g. water temperature) in the oceans as well as primary productivity (evaluated through the study of opal and phosphorus). Microfossils occur in such vast numbers that statistically significant samples can be collected systematically, unlike the vast majority of vertebrate fossil assemblages on land* where we have single specimens or only a handful of a particular taxon in question.
*I still think much more can be done with shark teeth in continental shelf marine rocks than just ‘here’s a list of the species we found in this layer’, but I digress.
Elizabeth Sibert’s prior projects including with her mentor Dick Norris* have focused on the K/Pg extinction in sharks and fish and found that sharks became numerically less common right after the extinction, previously outnumbering fish teeth, but after the extinction fish teeth become more common than shark denticles (Sibert and Norris, 2015). In another study she found that pelagic fish “ichthyoliths” (fish teeth in deep sea sediments) become numerically less common in the Tethys Sea for 3-4 million years after the K/Pg extinction but in the mid-Pacific, fish “ichthyoliths” remain constant and suggest only minor perturbation of the food chain, indicating that marine ecosystems were not uniformly affected by the extinction despite loss of primary producers like zooplankton (Sibert et al., 2014). A study of fish teeth and shark denticles from core samples from across the North and South Pacific found three major different long-term faunas from the past 85 million years: 1) A Cretaceous fauna (85-65 myo) dominated by shark denticles, 2) a Paleogene fauna (65-20 myo) where sharks were slightly less common than fish but both were relatively abundant and 3) the modern fauna (20-0 myo) where denticles are very rare, perhaps 1 for every 50 fish teeth, but also smaller amounts of ichthyoliths that are more variable in vertical distribution within the core samples (Sibert et al., 2016). A useful overview of the methods developed for these studies (and modified from micropaleontology) is provided by Sibert et al. (2017).
One caveat is that shark teeth are generally considered much more diagnostic and the majority of the shark fossil record and taxonomy of extinct sharks has been based on isolated teeth – not denticles. In this regard, denticles are used as a bit of a rough proxy for diversity – and quite a bit rougher than if only teeth were used. More on this below.

Methods and Marine Vertebrate Taphonomy
So how do you study ‘ichthyoliths’? They need to be removed from the sediment within the core sample. In order to facilitate removal, the sample sediment is typically rinsed in dilute acetic acid (5% or 10%) unless preserved in clay, in which case it is just rinsed in water. After this, the sediment is rinsed in deionized water and or bleach; staining may be needed to make picking easier. “Ichthyoliths” are then picked one by one from the disaggregated sediment samples. Floating the fossils using heavy liquid (e.g. kerosene) might be needed in some cases to separate them from other non-fossil grains. The use of acid means that no foraminifera or other calcareous microfossils will be recovered since they will be completely dissolved. After picking, they are identified to any number of morphotypes; in the case of the Sibert and Rubin (2021A) study, they assigned denticles to 85 different morphotypes. Many of these are not diagnostic and cannot be identified to a particular genus – many sharks have similar denticles and multiple denticle types (we will come back to this in gory detail) and most fish have little conical teeth. But some sense of the diversity can be gleaned by dividing these into purely descriptive morphotypes that are generally free of judgemental taxonomic baggage. Once identified, you can count the number per morphotype – or just the number of shark denticles or fish teeth altogether – per vertical measurement (e.g. centimeters) of the core sample. Age models are produced by analyzing strontium isotope ratios, matching to the global strontium curve, and estimating sedimentation rate for the remainder of the core sample – generally thought to be slow but continuous in the deep sea, however sedimentation rate can slow down or cease completely. Once ages are assigned to the different parts of the column, an “accumulation rate” for the fossils can be assessed – X number of teeth/denticles deposited in some volume of rock over a time period. For example, denticles were being deposited at a rate of 10-30 denticles per cm^(-2) per year about 20 million years ago, just before the purported extinction. More discussion of the micropaleontological methods adapted for vertebrate microfossils (ichthyoliths) were presented in Sibert et al. (2017).
*Norris was a student of prominent marine taphonomist Susan Kidwell and his Master’s thesis research focused on the fossil invertebrates of the Purisima Formation near Santa Cruz and Half Moon Bay, California – his study, published in 1986, was a major starting point for my own work on the vertebrate taphonomy of the unit which also sought to reconcile Purisima sedimentology and stratigraphy within a sequence stratigraphic framework. Norris has since been working at Scripps in La Jolla on Paleoceanographic research based on studies of deep sea plankton from core samples.
Now – some comments on taphonomy before we continue – there is an inverse relationship between sedimentation rate* and the abundance of vertebrate fossils. Vertebrate fossils are generally rare compared to invertebrates like mollusks and plankton in marine deposits; vertebrate fossils only seem to be common in strata deposited during periods of slow sedimentation rate. When sedimentation rate slows down, there are more vertebrate fossils per unit volume of sediment. This is why at fast sedimentation rates, vertebrate fossils are frequently so rare that they are nearly impossible to find during fieldwork. Only assemblages that were deposited during slow sedimentation rates are worth targeting for quarrying or screening, and include marine bonebeds. Another issue here is that in most marine strata, vertebrate fossils are so rare outside bonebeds that accumulation rates – how many teeth/denticles/bones per unit volume of sediment/year are fossilized in the strata – cannot be calculated, so we have generally assumed a constant accumulation rate for vertebrates. Studies of marine invertebrates, largely by Susan Kidwell, demonstrated that changes in sedimentation rate AND mollusk accumulation rate can both change. For the purposes of my taphonomic research, the lack of such a high resolution age model and rarity of vertebrate specimens outside bonebeds meant such an approach was not really feasible. But, those problems aren’t an issue with deep sea core samples. To summarize: in marine vertebrate taphonomy we usually assume that increases in the abundance of vertebrate fossils correlates most strongly with slow-downs in sedimentation rate. Bonebeds rich in teeth and bones form when sedimentation slows down or pauses (hiatal concentrations – e.g. formed during a sedimentary hiatus, or a condensed section) and lag deposits can form when erosion scours away sediment and leaves the heavier fossils behind (erosion is a ‘negative’ sedimentation rate).
*Sedimentation rate is simply measuring the vertical thickness of sediments deposited per unit time, usually in millions of years. Deposition rate is something else that avoids the problem of non-deposition, but it’s been 15 years since I took my last stratigraphy class and can’t remember precisely. Whatever it was it was such an esoteric argument that a prolonged google search is failing me – in any event, my advanced stratigraphy professor was a bit of a dick who had an axe to grind against sequence stratigraphy and the argument might have just been one of his ‘pet’ arguments anyway (of which he had many which were ultimately meaningless in the contemporary landscape of sedimentary geology).

Findings of Sibert and Rubin (2021A)
The study by Sibert and Rubin, published in Science, found several lines of evidence suggesting that an extinction or something similar affected pelagic shark and fish communities. Recall already that prior studies by Elizabeth Sibert had found evidence of a major discontinuity in the shark/fish ichthyolith record right about 20 myo. The first line of evidence is that many of the denticle morphotypes disappear right around 19 myo – many other morphotypes disappear earlier, with notable terminations of multiple morphotypes during the middle Oligocene at 30 myo, the Eocene-Oligocene boundary at 34 myo (associated with the onset of Antarctic glaciation), and at the Oligocene-Miocene boundary at 23 myo – if their figure 1 is interpreted literally. Instead, these authors suggest that many of these lineages disappear at the 19 myo mark and some earlier terminations might be better explained by signor-lipps effect, which suggests that at “low” sample sizes in the fossil record, a mass extinction event might be smeared out, with many species appearing to go extinct before the extinction event owing to the vagaries of fossil preservation. According to these authors, around 70% of denticle morphotype diversity disappears during the early Miocene, which is higher than the extinction rate (~40%) at the K/Pg boundary based on prior studies by Sibert using the same methods. Of the two broad denticle types – linear, characterized by parallel ridges, and geometric, characterized by more complicated shapes – both suffered considerably (66% and 88% extinction of morphotypes, respectively) but geometric denticle types declined in abundance, from 35% of denticles prior to 19 myo to only 3% of all denticles after 19 myo. Most sharks today have linear denticles, and geometric denticles are only present in small deep sea squaliform sharks like the lantern shark Etmopterus and the cookiecutter shark Isistius.

As a result of these findings, Sibert and Rubin interpreted this to indicate an extinction that selectively affected deep sea or pelagic sharks. These authors do point out that there is a considerable hiatus in sedimentation at many deep sea drilling sites, and few core samples have a continuous record through the early Miocene. Another issue, explored below, is that extant sharks exhibit a multitude of denticle types even within a single individual shark, and it would be a pretty monumental task to survey all denticle types of all extant shark species. Sibert and Rubin considered most modern shark species to only exhibit 1 or 2 denticle morphotypes, and they surveyed 152 species – which is a daunting task, no doubt, but a small proportion of extant species. It is conceivable, even if every family is represented in their extant denticle catalog, that some morphotypes might be missed altogether given that there are some 1,000+ species of elasmobranchs and about 27% of them were studied (27% according to Feichtinger et al.).
Challenges to the Early Miocene Extinction Hypothesis: rebuttals by Feichtinger et al. & Naylor et al.
Within a few months of publication, two rebuttals by two different teams of ichthyologists and paleoichthyologists were published – the first is largely a paleontological response by Iris Feichtinger et al. with many well-known paleoichthyologists, including Sylvain Adnet, Gilles Cuny, Jurgen Kriwet, Juergen Pollerspock, Kenshu Shimada, Charlie Underwood, Romain Vullo, and others. The second rebuttal is by Gavin Naylor and colleagues (including famous collector and shark expert Gordon Hubbell) – Naylor, by the way, worked here at CofC before leaving for greener pastures a few years ago – and this study is largely ichthyological in nature.

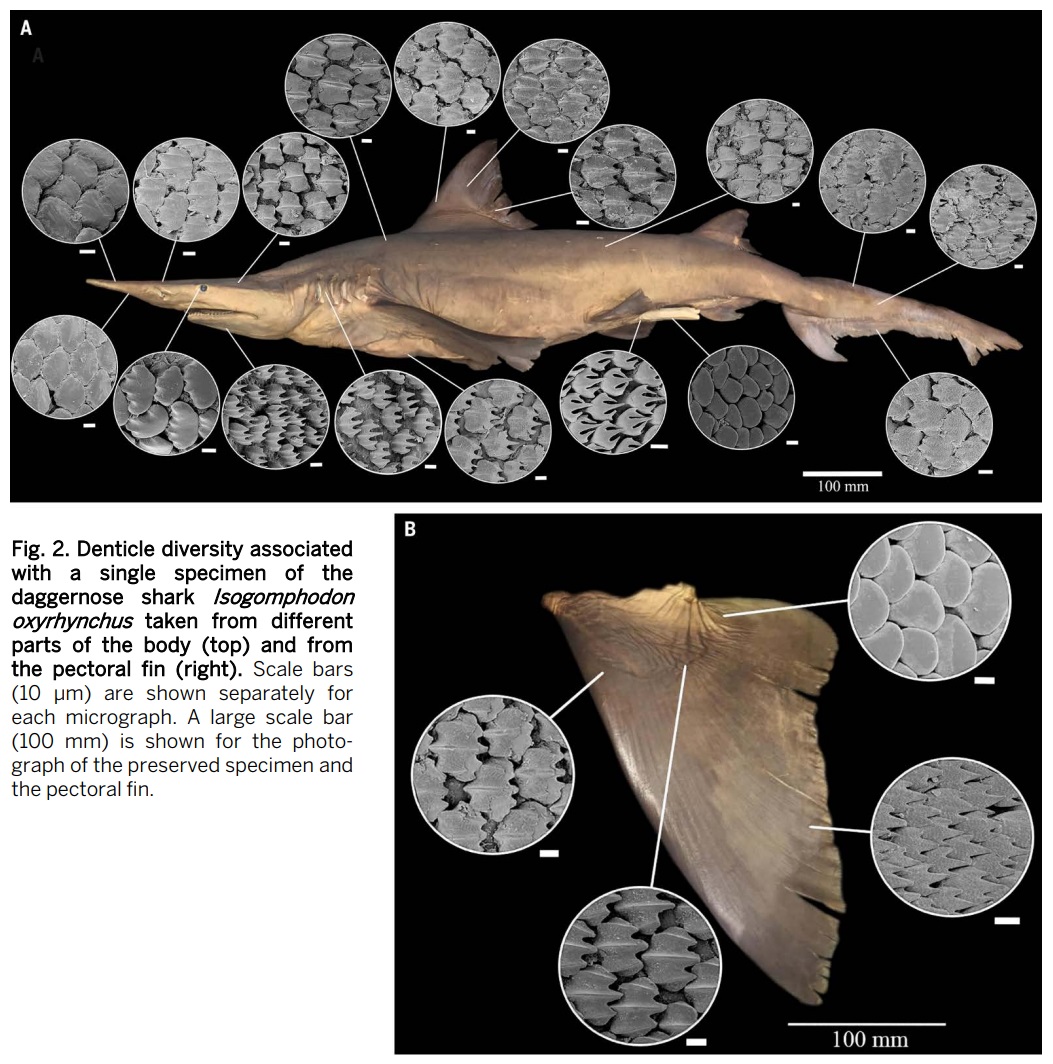
One major concern issued by both studies, and accompanied by some pretty amazing figures, is the alleged underestimate of denticle types by Sibert and Rubin – each of the two rebuttals notes that many denticle types can be present in a single individual shark; Feichtinger et al. show the denticles of a lantern shark, Etmopterus, and Naylor et al. show regional differences in the denticles of a daggernose shark, Isogomphodon. Each figure is intended to showcase differences in denticle morphology across the body of one individual shark – but no attempt was made by either set of authors to try to assign denticles from their individual sharks to the morphotypes of Sibert and Rubin. There are some differences in the denticles of Feichtinger’s Etmopterus, but I can imagine Sibert and Rubin may have considered these to be lumped into 2-3 ‘geometric’ morphotypes. However, this is admittedly much less possible with the Isogomphodon, which, from my mostly ignorant perspective, would have at least 5-6 ‘linear’ morphotypes. Regardless, had I been a reviewer of either rebuttal, I would have expected Feichtinger et al. or Naylor et al. to replicate the methods and assign denticles of a single individual modern shark to the morphotypes; neither set of authors can hardly claim the method is opaque, as Sibert and Rubin have a very detailed appendix with example denticles for each morphotype well-illustrated. Feichtinger et al. claim that nearly all of the denticles that disappear around 19 myo could have belonged to a single shark species, and just a single species going extinct could produce the pattern of denticle extinction. Naylor et al. make a similar argument, that the regional extirpation of just a few species of deep sea sharks could have produced the same pattern.

Feichtinger et al. also make the point that the purported extinction event corresponds to a major discontinuity in sedimentation rate – with a major increase in sedimentation rate beginning around 19 myo, and likely explaining the lesser abundance of fossils. They also point out a decrease in the abundance of bony fish fossils during this interval, which Sibert and Rubin did not dwell on. Feichtinger et al. argue that the much lower sample size after 19 myo in the samples resulted in the difference in the shark denticle:fish teeth ratio and is best explained as a sampling artifact. When focusing only on samples that could reasonably be expected to include at least one denticle – those having at least 6 fish teeth – only a few samples are informative.
Naylor et al. also make a claim which I find to be in ignorance of the fossil record – it’s a fair claim to be made by a biologist, but one not founded in paleontological familiarity. They argue that the lack of teeth – and dominance of denticles in the core samples – means the samples are from a “biased depositional environment-one that evidently favors the preservation of bony fish teeth and shark denticles, but strangely, not shark’s teeth.” This is a bit of a problem, as teeth are far more diagnostic than denticles. I have no data on the relative abundance of shark teeth and denticles, but even in tooth-rich stratigraphically thin (and therefore time-rich) bonebeds, shark teeth are rarely ever more common than a few miniscule teeth per kilogram of sediment. The drills used for Sibert & Rubin’s deep sea sampling are approximately 6 cm in diameter (~4.5 inches), but hitting a shark tooth with a drill in deep sea sediments would be like finding the proverbial needle in the haystack. Sharks famously produce and shed thousands of teeth in their lifespan – 10,000 is a commonly found number on the web – but have many thousands of denticles for each shark. The denticles are shed just like teeth (and our skin) and must be several orders of magnitude more common in deep sea sediments as a result. And of course, let us return to the sample size of the original Sibert and Rubin study: a total of 1263 denticles from two different core samples. Even if we assume that sharks ONLY have 1,000 denticles for every tooth, which I think is probably still being extremely generous, we’d expect a single tooth to have been found for this sample*. In sum, Naylor et al. either made this argument in bad faith (e.g. the tooth:denticle ratio in sharks) or are just not aware of the realities of the shark fossil record: from a tooth/sediment volume perspective, shark teeth are extremely rare fossils even when they are concentrated into tooth-rich bonebeds. Vertebrate macrofossils are rarely encountered in core samples, to the point that micropaleontologists usually treat them as oddities; I can think of only two mammal fossils, for example, ever found in core samples – one was a pinniped tooth in a Holocene-Pleistocene core sample from Antarctica someone emailed me about around a decade ago, and another is a ?Paleocene mammal in a core sample from Louisiana I recently learned about on Twitter.
*Sibert and Rubin responded to this (see below) and I wrote this prior to re-reading their response! Glad to know I was right on the money.
Returning to shark teeth – Naylor et al. point out that no extinctions are known in fossil shark assemblages during the early Miocene, and that all known shark families persist through the 19 my interval. They do allow however that undocumented pelagic shark taxa might be affected by this event – the implication being that they may not be expected to be found in continental shelf rocks. Truth be told, most continental shelf marine deposits were deposited at shallow or mid shelf depths over the past 30 million years, and contain sharks that are largely shallow marine – many of which still live on our shelves. Where do you need to go to even find deep-sea shark fossils? I cannot think of any on the east coast, aside from a handful of cookiecutter shark teeth (Isistius) from a single creek in Florida – and on the west coast, there is only a few Neogene deep sea shark assemblages, like the Skooner Gulch Formation in Mendocino County. Others include mid-Cenozoic assemblages in the Pacific Northwest like the Makah and Pysht formations of Washington, more famous for their early marine mammal fossils. One critical issue is that in these deep-sea deposits, shark teeth are only rarely if ever found at the outcrop, owing to rapid and continuous sedimentation rates. Many are found during the preparation of other marine vertebrates – I recently found something like an Etmopterus tooth while acid-prepping an early cetacean skull from the Lincoln Creek Formation.
There are no shortage of deep marine shark assemblages from the Mesozoic, BUT this is because these rocks are older and have had plenty of time to be uplifted: the same cannot really be said for Miocene assemblages even on active margins on the Pacific coast, where deep sea deposits are frequently still buried. On a passive continental margin like the Atlantic coast, the continental shelf has never been deeply immersed enough for deep sea sharks to shed their denticles and teeth into sediments that we can collect from. This in a way is a backwards ‘pull of the recent’ – our terrestrial fossil record gets better and better the closer we get to the Pleistocene and Holocene, but our marine record gets terrible. Why is that? It’s because most of the marine rocks are still under the ocean, aside from only the shallowest deposits. I’ve made this argument before in comparing the marine mammal record of ‘lagerstatten’ with marine reptiles – there just aren’t that many deep marine deposits of laminated, black shales in comparison with the Mesozoic because these deposits are probably still under the ocean or way way out on the continental shelf (aside from the greater oxygenation of Cenozoic seas and lack of stratified, anoxic seas).
Sibert and Rubin respond to criticism from Feichtinger et al. and Naylor et al.
One of the most critical points brought up by Feichtinger et al. is the major change in sedimentation rate at approximately the same horizon as the inferred extinction. However, Sibert and Rubin point out that their accumulation rate normalizes/accounts for changes in sedimentation rate. In other words, the analysis should be robust to changes in sedimentation rate. The low sample sizes are perhaps still a problem, but accumulation rate is pretty widely used in micropaleontology. However, I’m not familiar enough with the method to comment further – though perhaps the same can be said for Feichtinger et al. As for the fish tooth to denticle ratio, Sibert and Rubin indicate that in well-sampled horizons, denticles are outnumbered by fish teeth by 1:37 to as much as 1:70, as opposed to a ratio of 1:5 before the purported extinction – suggestive of some biological phenomenon at minimum, and perhaps indicating that the rarity of denticles is not necessarily a sampling artifact. Rarefaction analyses by Sibert and Rubin (2021B) indicate that further sampling would not have changed the pattern of denticle extinction.

What about the concern addressed by both Naylor et al. and Feichtinger et al. that Sibert and Rubin underestimated denticle variation in sharks, and that the disappearance of just a couple of species (through a relatively minor extinction or extirpation) could explain the pattern of denticles without necessitating some ocean-wide pelagic extinction event. Sibert and Rubin admit that their catalog is not exhaustive, but assert that it is good enough: they also performed a second rarefaction analysis (Sibert and Rubin 2021C) indicating that additional sampling of modern denticles would not have resulted in the recognition of many more denticle morphotypes. This is to a degree based on some simulations, and I do not have the background to judge this. For the uninitiated – rarefaction is a type of analysis used in ecology to figure out how many species can be expected to be encountered based on the sample size, given that small sample sizes/limited number of observations will only produce a small number of species, and the more observations are made, the more species will be recorded. At some point, the number of observations will result in diminishing returns and the number of observations should be “good enough”, so to speak – which Sibert and Rubin (2021C) assert their analysis supports. I am skeptical, to be honest, given the very different assessment of denticle morphotypes present in a single shark species by Sibert and Rubin (2021A; 1-2 morphotypes per species) versus Feichtinger et al. and Naylor et al. (not quantified, but to my estimation at least 3-5 morphotypes in their figured examples, conservatively speaking). With an estimated 27% of modern species sampled, there is a LOT of territory left; many shark species sampled by Sibert and Rubin include only a few skin samples, and based on the Naylor et al. image of Isogomphodon, perhaps not enough anatomical regions of sharks were sampled by Sibert and Rubin. If the initial catalog underestimates morphotype diversity, might that not mean that the rarefaction analyses and simulations may be inadequate? Again, I don’t know enough of the ins and outs of these analyses to comment further.

In another analysis, Sibert and Rubin (2021C) simulated the relationship between denticle morphotype extinction rate and species extinction rate, and found that for 70% of morphotypes to go extinct, perhaps 90% of species/genera might have to go extinct (recall that 70% is the observed rate of extinction of denticle morphotypes about 19 myo in Sibert and Rubin’s (2021A) analyses. However, if I understand this correctly, this is all based on the integrity and exhaustiveness of the original catalog.
Sibert and Rubin point out* that denticles outnumber teeth in an individual shark by three to five orders of magnitude – in other words, denticles are 1,000 to 100,000 times more common. Sibert and Rubin similarly point out (as I did above) that out of a sample under 2,000 denticles, only 1-2 teeth might be expected to be found.
*I wrote the above section where I criticized the Naylor et al. comments about not finding teeth before I read the response by Sibert and Rubin and am pretty chuffed that I was right on the money.
Is the extinction real? Some corollary evidence from marine mammal paleontology – or coincidences?
Both of the replies by Feichtinger et al. and Naylor et al. assert, perhaps correctly, that there are no known extinctions of sharks that we know of in marine fossil assemblages found in continental shelf sediments (e.g. on land). I’d wager that, based on what I know of Neogene shark paleontology, there’s quite a bit of fossils out there that have not yet actually been published so there is a helluva ‘black box’ remaining to be opened up. There are some differences between well-known Oligocene shark assemblages and those from younger Miocene rocks, but the timing of transitions and/or extinctions remains poorly established, with notable exceptions like excellent studies of Oligo-Miocene change in the snaggletooth shark Hemipristis (Chandler et al. 2006) and the loss of cusplets in Carcharocles (Perez et al., 2019). Lastly, don’t even get me started on Oligocene sharks: very, very few Oligocene assemblages are published so any claim that they’re well-studied is laughable.
Well, that’s enough about sharks: what about marine mammals? A couple major things happen during the early Miocene right around the time that the proposed extinction takes place. The first, and best established, is the widespread distribution of phocid seals throughout the Mediterranean and North Atlantic. Many of the oldest phocid seals are Burdigalian in age and Aquitanian specimens are rare or have dubious/poorly established ages. In the Pacific, pinnipeds diversify in the Burdigalian, with a sudden appearance of the first otariid fur seals like Eotaria and the first imagotariine walruses, most of the earliest desmatophocid seals, and some of the last-surviving enaliarctine ‘proto-seals’. The second event are changes in odontocete fossil assemblages, with the first common occurrences of early delphinoids like the typically small-bodied porpoise-like kentriodontids, which about ten million years later would give rise to the first belugas, true porpoises, and oceanic dolphins. In addition, sperm whales become common and arguably more diverse in the Burdigalian – including many large macrophagous species - though it is unclear how sudden or ‘blurry’ this was. The third, and perhaps most important and enigmatic, is the global rarity or outright absence of baleen whales in any earliest Miocene (23-20 myo, Aquitanian stage) deposits as documented by Marx et al. (2019). There are tons of toothed mysticetes, eomysticetids, and Mauicetus-grade whales (NZ only) in late Oligocene strata of the Pacific Northwest, Japan, Australia, New Zealand, and here in South Carolina – and pretty much only dolphins in Aquitanian-correlative deposits in the Pacific Northwest, Chesapeake Group in Maryland/Virginia, and the well-sampled Belluno Sandstones of Italy. Then, in the Burdigalian, the mysticete fossil record picks back up with a record dominated by numerous “kelloggitheres” – a moniker for Parietobalaena-grade early crown mysticetes which Remington Kellogg, the father of paleocetology, devoted no small time to studying as skeleton after skeleton kept on being discovered at Calvert Cliffs in Maryland. Marx et al. (2019) suggested that perhaps chaeomysticetes (toothless mysticetes) were predominantly offshore/pelagic during the Aquitanian, perhaps explaining their rarity in continental shelf strata. During the Aquitanian, there was a drop in sea level worldwide as well as a substantial decrease in the abundance of diatoms.
Changes in the pinniped fauna as well as the odontocete fauna correspond in a rough sense, but the sudden and widespread appearance of baleen whales after the Aquitanian I feel is perhaps more profound given the feeding ecology of baleen whales – they’re tied to prey low in the food chain (zooplankton). In my opinion, there’s plenty we do *not* know about the early Miocene, and there are just enough coincidences with faunal change in the marine mammal record to suggest that something happened in pelagic ecosystems.
Closing Thoughts
Altogether, I think this was a pretty fascinating back and forth over deep sea denticles. I’m not fully convinced that an extinction occurred – but I’m not convinced that it’s not “nothing”, either, and there’s still a lot we don’t know about marine vertebrate faunal evolution across the Oligocene-Miocene transition. I think further sampling is needed – both of deep sea denticles and modern shark denticles – and these remain pretty serious issues for the Sibert and Rubin analyses. In my opinion, the problem of undersampled extant shark denticle morphotypes is the single biggest issue with the analysis, and I am less concerned with sedimentation rate v. accumulation rate, as that strikes me as something that micropaleontologists sorted out decades ago. On a parting note, I’m really impressed by the sorts of groundbreaking work by Elizabeth Sibert and colleagues and am eagerly awaiting her next scientific contribution – it’s a new way of studying the shark and fish fossil record and it’s already made ‘big waves’ (hah hah) in marine vertebrate paleontology, compared to the alternative way we’ve been studying assemblages (one new locality at a time). Who knows what further sampling of modern shark denticles will result in!
References
Boessenecker and Churchill, 2018. https://academic.oup.com/zoolinnean/article/184/1/211/4783131
Feichtinger et al., 2021. https://www.science.org/doi/10.1126/science.abk0632
Marx et al., 2019. https://www.app.pan.pl/article/item/app005752018.html
Naylor et al, 2021. https://www.science.org/doi/10.1126/science.abj8723
Sibert et al., 2014. https://www.nature.com/articles/ngeo2227
Sibert and Norris, 2015. New Age of Fishes initiated by the Cretaceous−Paleogene mass extinction | PNAS
Sibert and Rubin, 2021A. https://www.science.org/doi/10.1126/science.aaz3549
Sibert and Rubin, 2021B – response to Feichtinger et al. 2021. https://www.science.org/doi/10.1126/science.abk1733
Sibert and Rubin, 2021C – response to Naylor et al. https://www.science.org/doi/10.1126/science.abj8723
Sibert et al., 2017. https://palaeo-electronica.org/content/2017/1800-quantifying-ichthyoliths
No comments:
Post a Comment