
Tohoraata - dawn whales from the Oligocene of New Zealand
Mauicetus waitakiensis was described in 1956 by former Zoology professor (and Zoology Chair) Brian J. Marples, although collected somewhat earlier by him. Marples was an arachnologist originally, and according to Ewan Fordyce, probably stumbled upon fossils while looking for trap door spiders in North Otago. During the 1940's and 1950's Marples and his sons dug up several cetacean skulls and partial skeletons, also including the dolphin Otekaikea marplesi, recently redescribed and renamed by my labmate/office partner Yoshi Tanaka. Two of these new cetaceans appeared to be baleen whales, and included one specimen with a complete braincase, fragmentary mandible, well-preserved tympanic bullae, scapulae, and some vertebrae; the other included only a fragmentary braincase, tympanic bullae, and neck vertebrae. Both were collected from the Kokoamu Greensand. At the time of writing, only one baleen whale had been named from NZ: Mauicetus parki, an archaic mysticete from the somewhat younger (earliest Miocene) Milburn Limestone, much closer to Otago campus (perhaps only a 45 minute drive out of town). At the time it was reasonable to allocate these new species to Mauicetus: the Mauicetus holotype was fragmentary, seemed to correspond well to what Marples had collected, and the periotic (inner ear bone) had not yet been removed. So, Marples named these two species Mauicetus lophocephalus (for the more complete specimen) and Mauicetus waitakiensis (for the less complete specimen).
Note: this can get somewhat confusing, as Dr. Benham originally named the type species as Lophocephalus parki (which he thought was an archaeocete, until Remington Kellogg corrected him), and upon realizing that Lophocephalus was preoccupied, he named the new genus Mauicetus for it. So Marples, perhaps confusingly, recycled the name for one of his new species of Mauicetus.
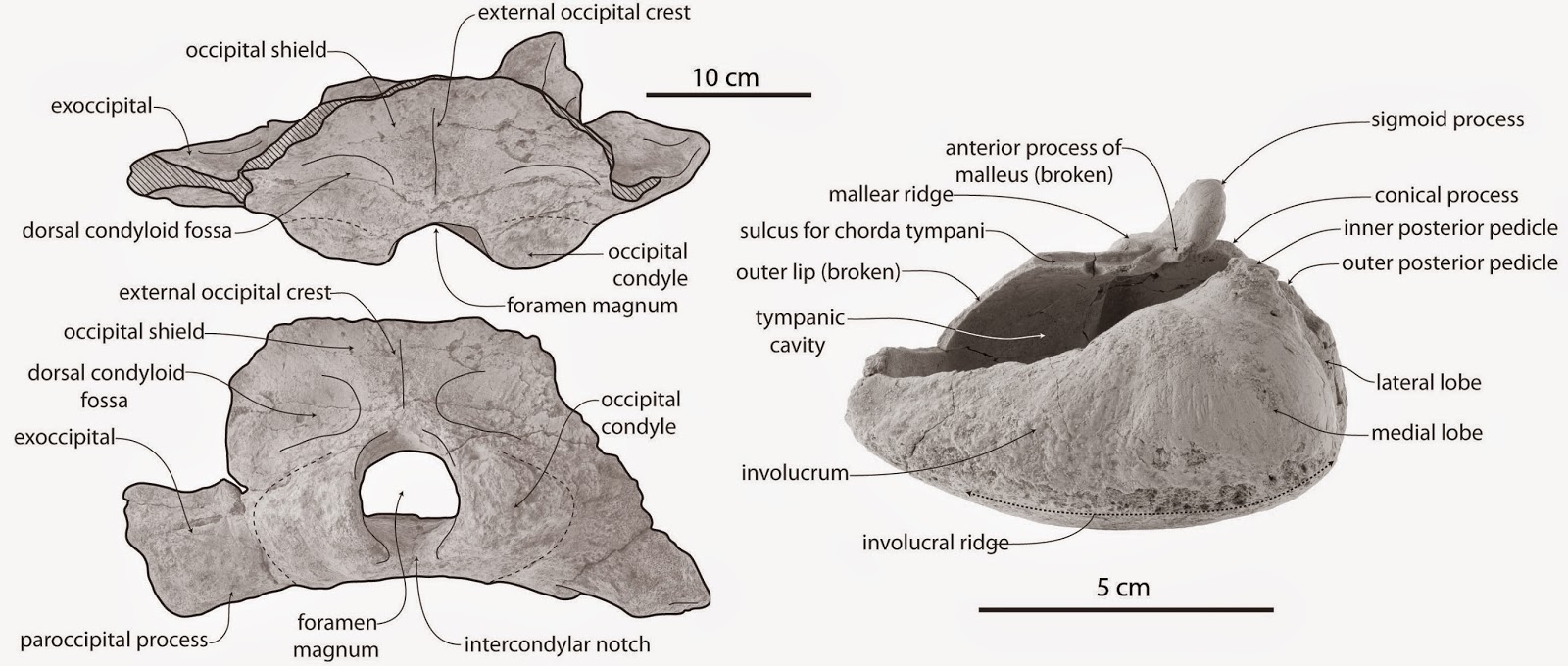
The holotype skull and tympanic bulla of "Mauicetus" waitakiensis, figured and described for the first time. From Boessenecker and Fordyce (2014A).
The relationships of Marples' Mauicetus species have remained contentious for nearly 60 years. Unfortunately, Marples never figured more than the first and second vertebrae of Mauicetus waitakiensis, even though it had a partial skull and earbones. To make matters worse, the skull of Mauicetus lophocephalus was chucked in the trash when the Zoology Dept. switched buildings sometime before 1962 - less than six years after being published! The next paleocetologist who came along and had the opportunity to look at this specimen was R.E. Fordyce when he began his Ph.D. in the late 1970's, and the skull of one of the most significant cetacean fossil discoveries in NZ had disappeared without a trace. Fordyce noted in several earlier studies that the remaining material of these two species differed strongly from all previously known baleen whales, and that the material likely represented a baleen-bearing mysticete but more primitive than any described. In 2002, Eomysticetus whitmorei was described by Al Sanders and Larry Barnes from the Oligocene Chandler Bridge Formation of South Carolina - an early toothless baleen whale that had features intermediate between toothed baleen whales and most groups of baleen whales from Neogene rocks, filling a critical gap in cetacean evolution. Eomysticetus had a long, narrow rostrum, a toothless palate, elongate nasal bones, an anteriorly placed blowhole, and an archaeocete-like braincase with archaeocete-like earbones and many archaeocete-like features of the postcrania (note: when I refer to archaeocetes, I'm specifically meaning advanced archaeocetes like basilosaurids). Ewan quickly realized that these specimens of Marples - including many unpublished fossils in our collections - represented similar species, likely referable to the family Eomysticetidae.

The holotype skull, bulla, and periotic of Tohoraata raekohao (OU 22178) from the upper Oligocene Otekaike Limestone, North Otago.
In 1993, Ewan and others collected another fragmentary mysticete from Waipati hill, the type locality of Waipatia maerewhenua (which would be formally named the following year). Other eomysticetid specimens in our collections are far more impressive - but shortly after my arrival here, this struck me as sharing the most in common with Marple's M. waitakiensis. This specimen included a partial skull, mandible, a single vertebra and several ribs, and most critically, well-preserved earbones. A few shared features (synapomorphies) of the tympanic bulla and occipital bone permitted these specimens to be identified, which we interpreted as reflecting a sister-species relationship within a single genus. We named this new genus Tohoraata, which in Maori literally translates as "dawn whale". The new species was named Tohoraata raekohao, which translates as "holes in forehead", referring to a series of peculiar foramina in the frontal bone above the eye socket (orbit); Marples' species was recombined as Tohoraata waitakiensis.

Yours truly posing with the displayed holotype specimen of Tohoraata raekohao. Photo copyright Otago Daily Times.
Discovery of the Tohoraata raekohao holotype not only represents the first record of an eomysticetid from the Oligocene of New Zealand, but also permitted taxonomic revision and identification of "Mauicetus" waitakiensis, resolving a half century of uncertainty over its relationships. This material also preserves the most exquisitely preserved tympanoperiotics of any published stem mysticete - and better yet, there are even better specimens from my thesis studies that have yet to be published (at least one far more spectacular fossil is the focus of another manuscript currently in review, including a revision of "Mauicetus" lophocephalus). Other fossils are more informative with respect to function, and will be featured in future publications.
Aside from publishing ammonium chloride coated photographs, we've constructed 3D models of the holotype periotic and bulla of Tohoraata raekohao, which are downloadable in pdf and obj format here.
Aside from publishing ammonium chloride coated photographs, we've constructed 3D models of the holotype periotic and bulla of Tohoraata raekohao, which are downloadable in pdf and obj format here.

A life restoration of Tohoraata raekohao with the skull digitally superimposed - copyright R.W. Boessenecker 2014.
Trace fossil evidence of fish/shark predation upon bone-eating worms
The second new publication also reports aspects of fossil eomysticetids from New Zealand, but is taphonomic in scope rather than taxonomic. Upon arriving down here I was surprised how many of the marine vertebrate fossils have circular pockmarks in them, measuring between 5 and 15 mm in diameter; in some cases, these pockmarks are coalesced and form large areas of bone where the entire surface has been destroyed. Similar pockmarks have been reported from Pliocene and modern whale bones, and in modern specimens, they have been confirmed to be made by the bone eating worm Osedax. Most previous studies of Osedax bioerosion have relied upon CT imaging - which is great, but expensive and impractical for the purposes of most taphonomic studies. Taphonomic studies hoping to mobilize large amounts of data cannot rely upon an expensive slow process such as non-invasive digital imaging, and rather ichnologic features useful for quick identification from a specimen will have more lasting significance for taphonomists. In other words, if you wanted to track the frequency of Osedax bioerosion through time - CT scanning hundreds of bones for a large quantitative study is totally impractical. Not that we had so many bones to work with for this project, we were dealing with a single specimen that had rather revealing traces - but it is a 2 meter long fossil baleen whale skull, and too large to scan at the necessary resolution (hospital CT scanners such as what we would have access to have a failsafe that scales the number of slices to the physical size of the object being scanned, so as to not expose a human patient to too much radiation - if I recall correctly). Also, we do not have the funding to pay for CT scans. So, we relied upon external characteristics in order to identify Osedax traces, which is standard practice anyway with virtually all ichnology (non-invasive digital imaging is generally done only in special cases).
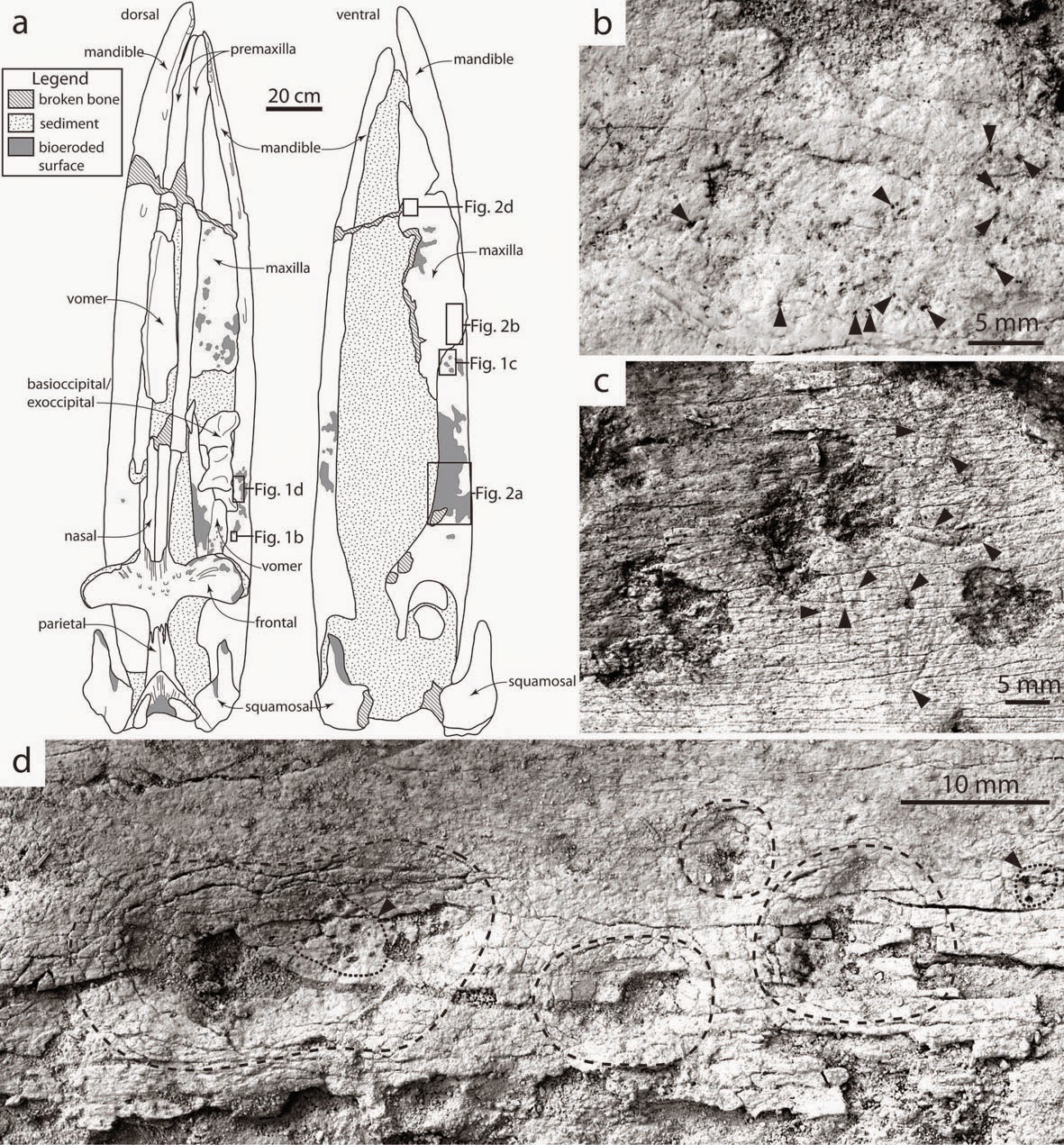
Illustration of the skull (A) of undescribed eomysticetid OU 22044 and various trace fossils found upon it. B: tiny boreholes from Osedax stalks; C, Osedax pockmarks with intersecting tooth scrapes; D, collapsed depressions with tiny boreholes.

More traces from OU 22044. A, bioeroded field of laterally coalesced Osedax pockmarks. B, set of three shark tooth scrapes. C, closeup of Osedax pockmarks with intersecting tooth scrapes. D, parallel set of tooth scrapes, likely from a skate.
This is of course neat, and represents the first record of fossil Osedax traces from the southern hemisphere - but they've been reported before and that novelty is not the selling point of the paper. Instead, the most significant discovery was that these tooth scrapes and pockmarks intersect - the scrapes cross-cut the damaged bioeroded pockmarks attributed to Osedax bioerosion. Osedax tends not to colonize bone surfaces until soft tissue is gone, although it has been observed colonizing soft tissues on sperm whale carcasses. Other tooth scrapes were likely made when soft tissue still remained, being inflicted during the process of scavenging. However, the scrapes that cross-cut the pockmarks are surficial, and only made after the pockmark already existed. If the bone was already colonized by Osedax, then presumably no soft tissue remained for the scavenger to consume. Instead, we attribute these scrapes as reflecting predation (by fish or shark) upon the Osedax worms themselves. Indeed, modern ratfish and tanner crabs have been observed gouging Osedax out of whale bones to consume the fleshy roots entombed within the bone. And now we have evidence for this behavior extending back to the Oligocene. On a broader philosophical level, we are reporting a predatory interaction between two organisms preserved only as traces in the skeletal tissues of a third organism (mind blown).
A couple other points of interest beg noting. 1) The existence of Osedax predators suggests that this process could accelerate the rate of skeletal bioerosion/recycling at the seafloor (if, of course, numerous phases of predation/recolonization happen). 2) We noted abundant pockmarks of similar morphology on fossils of bony fish, sea turtles, dolphins, baleen whales, and sea cows from Oligocene localities on the east coast of the USA as well as Europe, particularly from the Ashley/Chandler Bridge formations of South Carolina. In contrast, we noted no similar pockmarks on Neogene examples from either coast of the USA, including large assemblages from the Purisima, San Diego, San Mateo, and Calvert Formations as well as Sharktooth Hill. Perhaps widespread Osedax bioerosion (or preservation thereof) peaked in the Oligocene? We don't really know, and it's only a suggestion - but one that can be borne out by further (quantitative) study.
For more information, see our department web page (where you can download 3D models of the earbones!) on Tohoraata and these articles:
Stuff.co.nz: Ancient toothless whale skull discovered in Otago
Otago Daily Times: Otago researchers find new whale genus
3 News: 8m serpent like whale fossil found
New Zealand Herald: 20 million-year-old whale fossils found
Radio New Zealand: Researchers identify ancient NZ whale
Zee News: New Zealand researchers identify ancient whale species
Calgary Sun: N.Z. researchers uncover unknown ancient whales
Science World Report: Ancient New Zealand Whales Were the First to be Completely Toothless
Phys.org: Ancient New Zealand ‘Dawn Whale’ identified
These two papers are now out, and the first one on Tohoraata is even freely available online:
Boessenecker, R.W. and R.E. Fordyce. 2014A. A new eomysticetid (Mammalia: Cetacea) from the Late Oligocene of New Zealand and a re-evaluation of 'Mauicetus' waitakiensis. Papers in Palaeontology, Online early DOI:10.1002/spp2.1005
Boessenecker, R.W. and R.E. Fordyce. 2014B. Trace fossil evidence of predation upon bone-eating worms on a baleen whale skeleton from the Oligocene of New Zealand. Lethaia, Online early DOI: 10.1111/let.12108
No comments:
Post a Comment